Sekundaariset, tertiääriset, kvaternaariset proteiinirakenteet. Kemialliset sidokset, jotka osallistuvat proteiinirakenteiden muodostumiseen
P ERVICHNAYA RAKENNEBELKOV
Proteiinin primaarirakenne kantaa tietoa sen tilarakennetta.
1. Proteiinien peptidiketjun aminohappotähteet eivät vuorottele satunnaisesti, vaan ne on järjestetty tiettyyn järjestykseen. Polypeptidiketjun aminohappotähteiden lineaarista sekvenssiä kutsutaan proteiinin ensisijainen rakenne.
2. Kunkin yksittäisen proteiinin primäärirakenne on koodattu DNA-molekyylissä (osio, jota kutsutaan geeniksi), ja se toteutetaan transkription (mRNA:n tietojen uudelleenkirjoittaminen) ja translaation (peptidiketjusynteesi) aikana.
3. Jokaisella ihmiskehon 50 000 yksittäisestä proteiinista on ainutlaatuinen tietylle yksittäiselle proteiinille primaarirakenne. Kaikissa yksittäisen proteiinin (esimerkiksi albumiinin) molekyyleissä on sama aminohappotähteiden vuorottelu, mikä erottaa albumiinin kaikista muista yksittäisistä proteiineista.
4. Peptidiketjun aminohappotähteiden sekvenssiä voidaan pitää
tietuelomake
jotain tietoa.
Tämä tieto sanelee pitkän lineaarisen peptidiketjun avaruudellisen laskostumisen kompaktimmaksi kolmiulotteiseksi rakenteeksi.
KONFORMAATIOBELKOV
1. Yksittäisten proteiinien lineaariset polypeptidiketjut saavat aminohappojen funktionaalisten ryhmien vuorovaikutuksen vuoksi tietyn spatiaalisen kolmiulotteisen rakenteen tai konformaation. Globulaarisissa proteiineissa on
kaksi päätyyppiä konformaatioita peptidiketjut: sekundaariset ja tertiaariset rakenteet.
TOISIJAINENRAKENNEBELKOV
2. Proteiinien toissijainen rakenne on spatiaalinen rakenne, joka johtuu peptidirungon funktionaalisten ryhmien välisistä vuorovaikutuksista. Tässä tapauksessa peptidiketju voi saada säännöllisiä rakenteita kaksi tyyppiä:os-spiraalit Ja p-rakenteet.
Riisi. 1.2. Proteiinin toissijainen rakenne on a-heliksi.
Os-spiraalissa vetysidoksia muodostuu karboksyyliryhmän happiatomin ja veden välille peptidirungon amidityppisuvun kautta 4 aminohapon kautta; aminohappotähteiden sivuketjut sijaitsevat heliksin reunalla, eivätkä ne osallistu vetysidosten muodostumiseen, jotka muodostavat sekundaarirakenteen (kuva 1.2).
Suuret määrät tai jäämät, joilla on samat hylkivät varaukset, estävät yut a-heliksin muodostumiseen.
Proliinitähde katkaisee a-heliksin rengasrakenteensa vuoksi ja vetysidoksen muodostamisen mahdottomuudesta johtuen vedyn puuttumisesta peptidiketjun typpiatomissa.
B-Rakenne muodostuu yhden polypeptidiketjun lineaaristen alueiden väliin muodostaen laskoksia tai eri polypeptidiketjujen väliin. Polypeptidiketjuja tai niiden osia voi muodostua rinnakkain(vuorovaikutteisten peptidiketjujen N- ja C-päät ovat samat) tai vastakkainen(Vuorovaikutteisten peptidiketjujen N- ja C-päät ovat vastakkaisiin suuntiin) p-rakenteet(Kuva 1.3).
SISÄÄN proteiineissa on myös alueita, joilla on epäsäännöllinen sekundaarinen rakenne, joita kutsutaan sotkuiset pallot, vaikka nämä rakenteet eivät muutu niin paljon proteiinimolekyylistä toiseen.
TERTIÄÄRINENRAKENNEBELKOV
3. Proteiinin tertiäärinen rakenne- Tämä on kolmiulotteinen spatiaalinen rakenne, joka muodostuu aminohapporadikaalien välisistä vuorovaikutuksista, jotka voivat sijaita huomattavan etäisyyden päässä toisistaan peptidiketjussa.
Riisi. 1.3. Anti-rinnakkais (beta-rakenne.)
.jpg)
Aminohappojen hydrofobisilla radikaaleilla on taipumus yhdistyä proteiinien pallorakenteessa ns. opas-rofobisia vuorovaikutuksia ja molekyylien väliset van der Waalsin voimat muodostaen tiheän hydrofobisen ytimen. Hydrofiiliset ionisoidut ja ionisoimattomat aminohapporadikaalit sijaitsevat pääasiassa proteiinin pinnalla ja määrittävät sen vesiliukoisuuden.
Hydrofiiliset aminohapot, jotka ovat hydrofobisen ytimen sisällä, voivat olla vuorovaikutuksessa toistensa kanssa käyttämällä ioninen Ja vetysidoksia(riisi. 1.4).
.jpg)
Riisi. 1.4. Sidostyypit, jotka syntyvät aminohapporadikaalien välille proteiinin tertiaarisen rakenteen muodostumisen aikana. 1 - ionisidos; 2 - vetysidos; 3 - hydrofobiset vuorovaikutukset; 4 - disulfidisidos.
|
|
.jpg)
Riisi. 1.5. Disulfidisidokset ihmisinsuliinin rakenteessa.
Ioni-, vety- ja hydrofobiset sidokset ovat heikkoja: niiden energia ylittää hieman molekyylien lämpöliikkeen energian huoneenlämpötilassa.
Proteiinin konformaatiota ylläpitää monien tällaisten heikkojen sidosten esiintyminen.
Proteiinien konformationaalinen labilisuus- Tämä on proteiinien kyky muuttaa pieniä konformaatioita joidenkin hajoamisen ja muiden heikkojen sidosten muodostumisen vuoksi.
Joidenkin proteiinien tertiäärinen rakenne on stabiloitunut disulfidisidokset, muodostuu kahden kysteiinitähteen SH-ryhmien vuorovaikutuksesta.
Useimmissa solunsisäisissä proteiineissa ei ole kovalenttisia disulfidisidoksia. Niiden läsnäolo on ominaista solun erittämille proteiineille, esimerkiksi disulfidisidoksia on insuliinin, immunoglobuliinien molekyyleissä.
Insuliini- proteiinihormoni, joka syntetisoituu haiman β-soluissa. Solut erittävät sitä vasteena veren glukoosipitoisuuden nousulle. Insuliinin rakenteessa on 2 disulfidisidosta, jotka yhdistävät 2 polypeptidi-A- ja B-ketjua, ja 1 disulfidisidos A-ketjun sisällä (kuva 1.5).
Proteiinien sekundaarirakenteen ominaisuudet vaikuttavat interradikaalisten vuorovaikutusten luonteeseen ja tertiääriseen rakenteeseen.
4. Monissa proteiineissa, joilla on erilainen rakenne ja toiminta, havaitaan tietty sekundäärirakenteiden vaihtelujärjestys, ja sitä kutsutaan supersekundaarirakenteeksi.
Sellainen järjestettyjä rakenteita kutsutaan usein rakennemotiiveiksi, joilla on erityiset nimet: "helix-turn-a-helix", "leusiinivetoketju", "sinkkisormet", "P-tynnyrirakenne" jne.
A-heliksien ja p-rakenteiden läsnäolon perusteella pallomaiset proteiinit voidaan jakaa 4 luokkaan:
1. Ensimmäiseen luokkaan kuuluvat proteiinit, jotka sisältävät vain a-heliksiä, kuten myoglobiini ja hemoglobiini (kuva 1.6).
2. Toiseen kategoriaan kuuluvat proteiinit, joissa on a-kierteitä ja (3-rakenteita. Tässä tapauksessa a- ja (3-rakenteet) muodostavat usein samantyyppisiä yhdistelmiä, joita esiintyy eri yksittäisissä proteiineissa.
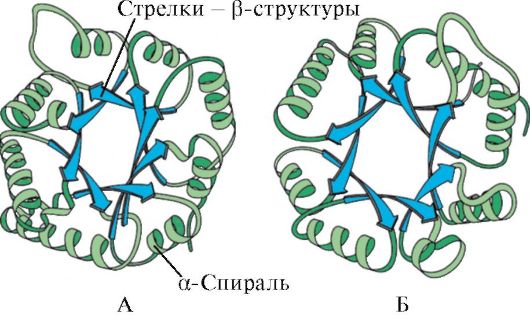
Esimerkki. Supersekundaarinen P-tynnyrirakenne.

Trioosifosfaatti-isomeraasientsyymillä on P-tynnyrityyppinen supersekundaarinen rakenne, jossa jokainen (3-rakenne sijaitsee p-tynnyrin sisällä ja liittyy polypeptidin a-kierteiseen alueeseenketjut sijaitsevat molekyylin pinnalla (kuva 1.7, A).
Riisi. 1.7. Supersekundaarinen p-tynnyrirakenne.
a - trioosifosfaatti-isomeraasi; b — Piru fleece nazan alue.
.jpg)
Sama supersekundaarinen rakenne löydettiin yhdestä pyrdomeenista (kuva 1.7b). Domeeni on osa molekyyliä, joka muistuttaa rakenteeltaan itsenäistä pallomaista proteiinia.
Toinen esimerkki supersekundaarisen rakenteen muodostamisesta P-rakenteiden ja os-heliksien kanssa. Yhdessä laktaattidehydrogenaasin (LDH) ja fosfoglyseraattikinaasin domeenissa polypeptidiketjun P-rakenteet sijaitsevat keskellä kierretyn levyn muodossa, ja jokainen p-rakenne liittyy a-kierteiseen alueeseen, joka sijaitsee molekyylin pinnalla (kuva 1.8).
Riisi. 1.8 Toissijainen rakenne, joka on ominainen monille fer- poliisit.
A- laktaattidehydrogenaasidomeeni; b- fosfoglyseraattikinaasidomeeni.
.jpg) 3. Kolmanteen luokkaan kuuluvat proteiinit, joilla on sisältää vain sekundaarisen p-rakenteen. Tällaisia rakenteita löytyy immunoglobuliineista, superoksididismutaasientsyymistä (kuva 1.9).
3. Kolmanteen luokkaan kuuluvat proteiinit, joilla on sisältää vain sekundaarisen p-rakenteen. Tällaisia rakenteita löytyy immunoglobuliineista, superoksididismutaasientsyymistä (kuva 1.9).
Riisi. 1.9. Immunoglobuliinin vakiodomeenin toissijainen rakenne (A)
ja entsyymi superoksididismutaasi (b).
.jpg)
4. Neljänteen kategoriaan kuuluvat proteiinit, jotka sisältävät vain pienen määrän säännöllisiä sekundaarisia rakenteita. Nämä proteiinit sisältävät pieniä kystiinirikkaita proteiineja tai metalloproteiineja.
DNA:ta sitovat proteiinit sisältävät yleisiä näkemyksiä supersekundaariset rakenteet: "os-spiral-turn-os-spiral", "leusiinivetoketju", "sinkki-sormesi." DNA:ta sitovat proteiinit sisältävät sitoutumiskohdan, joka on komplementaarinen DNA:n osalle, jolla on spesifinen nukleotidisekvenssi. Nämä proteiinit osallistuvat geenien toiminnan säätelyyn.
« A-
Spiraali-käännä-spiraali"
Riisi. 1.10. Sitova toissijainen
a-helix-turn-a-helix rakenteet
suuressa urassa D
|
|
.jpg)
.jpg) kaulan ura on hyvämukautettu sitomaan proteiineja ei-suurilla os-helikaalisilla alueilla.
kaulan ura on hyvämukautettu sitomaan proteiineja ei-suurilla os-helikaalisilla alueilla.
Tämä rakennemotiivi sisältää 2 os-heliksiä: yksi on lyhyempi, toinen on pidempi, yhdistettynä polypeptidiketjun käänteellä (kuva 1.10).
Lyhyempi a-heliksi sijaitsee DNA-uran poikki ja pidempi a-heliksi sijaitsee suuressa urassa muodostaen ei-kovalenttisia spesifisiä aminohapporadikaalien sidoksia DNA-nukleotidien kanssa.
Usein proteiinit, joilla on tällainen rakenne, muodostavat dimeerejä, minkä seurauksena oligomeerisellä proteiinilla on 2 supersekundaarista rakennetta.
Ne sijaitsevat tietyllä etäisyydellä toisistaan ja työntyvät proteiinin pinnan yläpuolelle (kuva 1.11).Kaksi tällaista rakennetta voi sitoutua DNA:han suurten urien vierekkäisillä alueilla.
ilmanmerkittäviä muutoksia proteiinien rakenteessa.
"sinkkisormi"
"Sinkkisormi" on proteiinifragmentti, joka sisältää noin 20 aminohappotähdettä (kuva 1.12).
Sinkkiatomi on kytketty 4 aminohapporadikaaliin: 2 kysteiinitähdettä ja 2 histidiinitähdettä.
Joissakin tapauksissa histidiinitähteiden sijasta on kysteiinijäämiä.
Riisi. 1.12. DNA:ta sitovan kohdan rakenneproteiineja "sinkkisormen" muodossa.
.jpg)
Tämä proteiinin alue muodostaa a-heliksin, joka voi spesifisesti sitoutua DNA:n pääuran säätelyalueisiin.
Yksittäisen säätely-DNA:ta sitovan proteiinin sitoutumisspesifisyys riippuu "sinkkisormen" -alueella sijaitsevien aminohappotähteiden sekvenssistä.
"Leusiini vetoketju"
Vuorovaikutuksessa olevilla proteiineilla on a-kierteinen alue, joka sisältää vähintään 4 leusiinitähdettä.
Leusiinitähteet sijaitsevat 6 aminohapon päässä toisistaan.
Koska jokainen a-heliksin kierros sisältää 3,6-aminohappotähteen, leusiiniradikaaleja löytyy joka toisen kierroksen pinnasta.
Yhden proteiinin a-heliksin leusiinitähteet voivat olla vuorovaikutuksessa toisen proteiinin leusiinitähteiden kanssa (hydrofobiset vuorovaikutukset), jotka yhdistävät ne toisiinsa (kuva 1.13).
.jpg) Monet DNA:ta sitovat proteiinit ovat vuorovaikutuksessa DNA:n kanssa oligomeeristen rakenteiden muodossa, joissa alayksiköt on liitetty toisiinsa "leusiinivetoketjuilla". Histonit voivat toimia esimerkkinä sellaisista proteiineista.
Monet DNA:ta sitovat proteiinit ovat vuorovaikutuksessa DNA:n kanssa oligomeeristen rakenteiden muodossa, joissa alayksiköt on liitetty toisiinsa "leusiinivetoketjuilla". Histonit voivat toimia esimerkkinä sellaisista proteiineista.
Histonit- ydinproteiinit, jotka sisältävät suuren määrän positiivisesti varautuneita aminohappoja - arginiini ja lysiini (jopa 80%).
Histonimolekyylit yhdistetään oligomeerisiksi komplekseiksi, jotka sisältävät 8 monomeeriä "leusiinikiinnittimien" avulla huolimatta näiden molekyylien voimakkaasta positiivisesta varauksesta.
Yhteenveto. Kaikki yksittäiset proteiinimolekyylit, joilla on identtinen primäärirakenne, saavat saman konformaation liuoksessa.
Täten, peptidiketjun spatiaalisen laskostumisen luonne määräytyy aminohapon perusteellaaminohappotähteiden koostumus ja vuorotteluketjut. Siksi konformaatio on aivan yhtä spesifinen yksittäisen proteiinin ominaisuus kuin primäärirakenne.
L yksittäisten proteiinien lineaariset polypeptidiketjut saavat aminohappojen funktionaalisten ryhmien vuorovaikutuksen vuoksi tietyn spatiaalisen kolmiulotteisen rakenteen, jota kutsutaan "konformaatioksi". Kaikki yksittäisten proteiinien molekyylit (eli joilla on sama primäärirakenne) muodostavat saman konformaation liuoksessa. Näin ollen kaikki tilarakenteiden muodostumiseen tarvittava tieto sijaitsee proteiinien primäärirakenteessa.
Proteiineissa on 2 päätyyppiä polypeptidiketjujen konformaatiota: sekundaariset ja tertiaariset rakenteet.
2. Proteiinien toissijainen rakenne - peptidirungon funktionaalisten ryhmien välisestä vuorovaikutuksesta johtuva spatiaalinen rakenne.
Tässä tapauksessa peptidiketjut voivat hankkia kahden tyyppisiä säännöllisiä rakenteita: α-heliksit
β-rakenneβ-rakenne ymmärretään hahmona, joka on samanlainen kuin haitariksi taitettu arkki. Kuvio muodostuu useiden vetysidosten muodostumisesta yhden polypeptidiketjun taivutuksia tekevien lineaaristen alueiden peptidiryhmien atomien välille tai eri polypeptidiryhmien välille.
Sidokset ovat vetyä, ne stabiloivat makromolekyylien erillisiä fragmentteja.
3. Proteiinien tertiäärinen rakenne - aminohapporadikaalien välisistä vuorovaikutuksista muodostuva kolmiulotteinen spatiaalinen rakenne, joka voi sijaita huomattavan etäisyyden päässä toisistaan polypeptidiketjussa.
Rakenteellisesti koostuu toissijaisen rakenteen elementeistä, stabiloitu erilaisia tyyppejä vuorovaikutuksia, joissa hydrofobisilla vuorovaikutuksilla on ratkaiseva rooli
proteiinin tertiaarisen rakenteen stabilointiin liittyy:
kovalenttiset sidokset (kahden kysteiinitähteen välillä - disulfidisillat);
ionisidokset aminohappotähteiden vastakkaisesti varautuneiden sivuryhmien välillä;
· vetysidokset;
hydrofiilis-hydrofobiset vuorovaikutukset. Kun proteiinimolekyyli on vuorovaikutuksessa ympäröivien vesimolekyylien kanssa, sillä "taipumus" käpertyä niin, että aminohappojen ei-polaariset sivuryhmät eristetään vesiliuoksesta; pooliset hydrofiiliset sivuryhmät ilmestyvät molekyylin pinnalle.
4. Kvaternaarirakennetta kutsutaan keskinäinen järjestely useita polypeptidiketjuja osana yhtä proteiinikompleksia. Proteiinimolekyylit, jotka muodostavat kvaternaarisen rakenteen omaavan proteiinin, muodostuvat erikseen ribosomeihin ja vasta synteesin päätyttyä muodostavat yhteisen supramolekyylirakenteen. Kvaternäärisen rakenteen omaava proteiini voi sisältää sekä identtisiä että erilaisia polypeptidiketjuja. Kvaternaarisen rakenteen stabilointiin osallistu samantyyppisiä vuorovaikutuksia kuin tertiaarisen stabiloinnissa. Supramolekulaariset proteiinikompleksit voivat koostua kymmenistä molekyyleistä.
Rooli.
Peptidien muodostuminen elimistössä tapahtuu muutamassa minuutissa, kun taas kemiallinen synteesi laboratoriossa on melko pitkä prosessi, joka voi kestää useita päiviä, ja synteesitekniikan kehittäminen kestää useita vuosia. Tästä huolimatta on kuitenkin olemassa melko painavia perusteita luonnollisten peptidien analogien synteesiä koskevan työn suorittamisen puolesta. Ensinnäkin modifioimalla kemiallisesti peptidejä on mahdollista vahvistaa primaarirakennehypoteesi. Joidenkin hormonien aminohapposekvenssit ovat tulleet tunnetuiksi juuri niiden analogien synteesin kautta laboratoriossa.
Toiseksi synteettiset peptidit antavat mahdollisuuden tutkia tarkemmin aminohapposekvenssin rakenteen ja sen aktiivisuuden välistä suhdetta. Peptidin spesifisen rakenteen ja sen biologisen aktiivisuuden välisen suhteen selvittämiseksi suoritettiin valtava määrä työtä yli tuhannen analogin synteesiä varten. Tuloksena oli mahdollista havaita, että vain yhden aminohapon korvaaminen peptidirakenteessa voi lisätä sen biologista aktiivisuutta useita kertoja tai muuttaa sen suuntaa. Aminohapposekvenssin pituuden muutos auttaa määrittämään peptidin aktiivisten keskusten sijainnin ja reseptorin vuorovaikutuskohdan.
Kolmanneksi, alkuperäisen aminohapposekvenssin muuttamisen ansiosta oli mahdollista saada farmakologisia valmisteita. Luonnollisten peptidien analogien luominen mahdollistaa molekyylien "tehokkaampien" konfiguraatioiden tunnistamisen, jotka tehostavat biologista vaikutusta tai pidentävät sitä.
Neljäs, kemiallinen synteesi peptidit ovat kustannustehokkaita. Useimmat terapeuttiset lääkkeet maksaisivat kymmenen kertaa enemmän, jos ne olisi valmistettu luonnontuotteesta.
Usein aktiivisia peptidejä löytyy luonnosta vain nanogrammamäärinä. Lisäksi menetelmät peptidien puhdistamiseksi ja eristämiseksi luonnollisia lähteitä ei voi täysin jakaa haluttua aminohapposekvenssiä vastakkaisen tai muun vaikutuksen omaavien peptidien kanssa. Ja ihmiskehon syntetisoimien spesifisten peptidien tapauksessa niitä voidaan saada vain synteesillä laboratoriossa.
57. Proteiinien luokittelu: yksinkertainen ja monimutkainen, pallomainen ja säikeinen, monomeerinen ja oligomeerinen. Proteiinien tehtävät kehossa.
Luokittelu rakennustyypin mukaan
Yleisen rakennetyypin mukaan proteiinit voidaan jakaa kolmeen ryhmään:
1. Fibrillaariset proteiinit - muodostavat polymeerejä, joiden rakenne on yleensä erittäin säännöllinen ja sitä tukevat pääasiassa eri ketjujen väliset vuorovaikutukset. Ne muodostavat mikrofilamentteja, mikrotubuluksia, fibrillejä, tukevat solujen ja kudosten rakennetta. Fibrillaarisia proteiineja ovat keratiini ja kollageeni.
2. Globulaariset proteiinit ovat vesiliukoisia, molekyylin yleinen muoto on enemmän tai vähemmän pallomainen.
3. Kalvoproteiinit - niillä on domeeneja, jotka läpäisevät solukalvon, mutta osa niistä työntyy kalvosta solujen väliseen ympäristöön ja solun sytoplasmaan. Kalvoproteiinit suorittavat reseptorien toiminnan, eli ne suorittavat signaalin välityksen ja tarjoavat myös kalvon läpi kulkevan kuljetuksen erilaisia aineita. Kuljettajaproteiinit ovat spesifisiä, jokainen niistä päästää vain tiettyjä molekyylejä tai tietyntyyppisen signaalin kulkemaan kalvon läpi.
Yksinkertaiset proteiinit , Monimutkaiset proteiinit
Peptidiketjujen lisäksi monet proteiinit sisältävät myös ei-aminohapporyhmiä, ja tämän kriteerin mukaan proteiinit jaetaan kahteen suureen ryhmään - yksinkertaisia ja monimutkaisia proteiineja(proteiinit). Yksinkertaiset proteiinit koostuvat vain polypeptidiketjuista, monimutkaiset proteiinit sisältävät myös ei-aminohapporyhmiä eli proteettisia ryhmiä.
Yksinkertainen.
Globulaaristen proteiinien joukossa ovat:
1. albumiinit - liukenee veteen laajalla pH-alueella (4 - 8,5), saostetaan 70-100 % ammoniumsulfaattiliuoksella;
2. polyfunktionaaliset globuliinit, joilla on suurempi molekyylipaino, vaikeammin liukenevat veteen, liukenevat suolaliuoksiin, sisältävät usein hiilihydraattiosan;
3. histonit - pienimolekyylipainoiset proteiinit korkea sisältö arginiini- ja lysiinitähteiden molekyylissä, mikä määrittää niiden perusominaisuudet;
4. protamiinit erottuvat vielä suuremmasta arginiinipitoisuudesta (jopa 85 %), kuten histonit, ne muodostavat stabiileja assosiaatioita nukleiinihappojen kanssa, toimivat säätely- ja repressoriproteiineina - olennainen osa nukleoproteiineja;
5. Proliineille on ominaista korkea glutamiinihapon (30-45 %) ja proliinin (jopa 15 %) pitoisuus, liukenematon veteen, liukenee 50-90 % etanoliin;
6. Gluteliinit sisältävät noin 45 % glutamiinihappoa, kuten prolamiinit, niitä löytyy useammin viljan proteiineista.
Fibrillaarisille proteiineille on tunnusomaista kuiturakenne, joka on käytännössä liukenematon veteen ja suolaliuoksiin. Molekyylien polypeptidiketjut on järjestetty rinnakkain toistensa kanssa. Osallistu sidekudoksen rakenneosien (kollageeni, keratiini, elastiini) muodostumiseen.
Monimutkaiset proteiinit
(proteiinit, holoproteiinit) - kaksikomponenttiset proteiinit, joissa peptidiketjujen (yksinkertainen proteiini) lisäksi sisältyy ei-aminohappoluonteinen komponentti - proteettinen ryhmä. Monimutkaisten proteiinien hydrolyysin aikana vapautuu aminohappojen lisäksi ei-proteiiniosa tai sen hajoamistuotteet.
Prosteettisena ryhmänä voivat toimia erilaiset orgaaniset (lipidit, hiilihydraatit) ja epäorgaaniset (metallit) aineet.
Prosteettisten ryhmien kemiallisesta luonteesta riippuen monimutkaisten proteiinien joukossa erotetaan seuraavat luokat:
· Prosteettisena ryhmänä kovalenttisesti sitoutuneita hiilihydraattijäännöksiä sisältävät glykoproteiinit ja niiden alaluokka - proteoglykaanit,ä. Seriinin tai treoniinin hydroksyyliryhmät osallistuvat yleensä sidosten muodostumiseen hiilihydraattitähteiden kanssa. Suurin osa solunulkoisista proteiineista, erityisesti immunoglobuliinit, ovat glykoproteiineja. Proteoglykaaneissa hiilihydraattiosa on ~95 %; ne ovat solunulkoisen matriisin pääkomponentti.
Lipoproteiinit, jotka sisältävät ei-kovalenttisesti sitoutuneita lipidejä proteettisena osana. Proteiinien muodostamat lipoproteiinit-apolipoproteiinit, joihin lipidit sitoutuvat ja suorittavat lipidien kuljetustoiminnon.
· Metalloproteiinit, jotka sisältävät ei-hemi-koordinoituja metalli-ioneja. Metalloproteiineista löytyy varastointi- ja kuljetustoimintoja suorittavia proteiineja (esim. rautaa sisältävä ferritiini ja transferriini) ja entsyymejä (esim. sinkkipitoinen hiilihappoanhydraasi ja erilaiset superoksididismutaasit, jotka sisältävät aktiivisina keskuksina kuparia, mangaania, rautaa ja muita metalli-ioneja )
Nukleoproteiinit, jotka sisältävät ei-kovalenttisesti sitoutunutta DNA:ta tai RNA:ta, erityisesti kromatiini, joka muodostaa kromosomeja, on nukleoproteiini.
Fosfoproteiinit, jotka sisältävät kovalenttisesti sitoutuneita fosforihappojäännöksiä proteettisena ryhmänä. Seriinin tai treoniinin hydroksyyliryhmät osallistuvat esterisidoksen muodostumiseen fosfaatin kanssa, fosfoproteiineja ovat erityisesti maidon kaseiini:
Kromoproteiinit - monimutkaisten proteiinien kollektiivinen nimi, joissa on värillisiä proteettisia ryhmiä, joilla on erilainen kemiallinen luonne. Näitä ovat monet proteiinit, joissa on metallia sisältävä porfyriiniproteesiryhmä, jotka suorittavat erilaisia toimintoja - hemoproteiinit (hemeä sisältävät proteiinit - hemoglobiini, sytokromit jne. proteettisena ryhmänä), klorofyllit; flavoproteiinit, joissa on flaviiniryhmä jne.
1. Rakenteellinen toiminta
2. Suojaustoiminto
3. Sääntelytoiminto
4. Hälytystoiminto
5. Kuljetustoiminto
6. Vara (vara)toiminto
7. Reseptoritoiminto
8. Moottorin (moottorin) toiminto
Proteiinit ovat polymeerisiä molekyylejä, joissa aminohapot toimivat monomeereinä. Ihmiskehon proteiinien koostumuksessa on vain 20 α-aminohappoa. Samat aminohapot ovat läsnä proteiineissa, joilla on erilaiset rakenteet ja toiminnot. Proteiinimolekyylien yksilöllisyys määräytyy proteiinin aminohappojen vuorottelujärjestyksen mukaan. Aminohappoja voidaan pitää aakkosten kirjaimina, joiden avulla, kuten sanalla, tiedot tallennetaan. Sana kantaa tietoa esimerkiksi kohteesta tai toiminnasta, ja proteiinin aminohapposekvenssi kantaa tietoa tämän proteiinin avaruudellisen rakenteen rakenteesta ja toiminnasta.
Aminohappojen yleinen rakenteellinen piirre on samaan a-hiiliatomiin liittyneiden amino- ja karboksyyliryhmien läsnäolo. R - aminohapporadikaali - yksinkertaisimmassa tapauksessa sitä edustaa vetyatomi (glysiini), mutta sillä voi olla myös monimutkaisempi rakenne.
Kaikki 20 ihmiskehon aminohappoa eroavat a-hiiliatomiin kiinnittyneiden radikaalien rakenteesta, koosta ja fysikaalis-kemiallisista ominaisuuksista.
Kemiallisen rakenteen mukaan aminohapot voidaan jakaa alifaattisiin, aromaattisiin ja heterosyklisiin (taulukot 1-1).

Aminohapot voivat sitoutua kovalenttisesti toisiinsa peptidisidosten kautta. Peptidisidos muodostuu yhden aminohapon a-karboksyyliryhmän ja toisen aminohapon a-aminoryhmän välille, ts. on amidisidos. Tässä tapauksessa vesimolekyyli hajoaa.
Peptidiketjut sisältävät kymmeniä, satoja ja tuhansia aminohappotähteitä, jotka on yhdistetty vahvoilla peptidisidoksilla. Molekyylisisäisten vuorovaikutusten vuoksi proteiinit muodostavat tietyn tilarakenteen, jota kutsutaan nimellä proteiinin konformaatio. Proteiinin lineaarinen aminohapposekvenssi sisältää tietoa kolmiulotteisen spatiaalisen rakenteen rakentamisesta. Proteiinien rakenteellisessa organisaatiossa on 4 tasoa, joita kutsutaan primäärisiksi, sekundaarisiksi, tertiäärisiksi ja kvaternaarisiksi rakenteiksi (kuvat 1-3). On olemassa yleisiä sääntöjä, joiden mukaan proteiinien spatiaaliset rakenteet muodostuvat.
Aminohappotähteet proteiinien peptidiketjussa eivät vuorottele satunnaisesti, vaan ne on järjestetty tiettyyn järjestykseen. Polypeptidiketjun aminohappotähteiden lineaarista sekvenssiä kutsutaan "proteiinin ensisijainen rakenne". Yksittäisten proteiinien lineaariset polypeptidiketjut saavat aminohappojen funktionaalisten ryhmien vuorovaikutuksen vuoksi tietyn tilallisen kolmiulotteisen rakenteen, ns. "muotoilu". Kaikki yksittäisten proteiinien molekyylit (eli joilla on sama primäärirakenne) muodostavat saman konformaation liuoksessa. Näin ollen kaikki tilarakenteiden muodostumiseen tarvittava tieto sijaitsee proteiinien primäärirakenteessa.
Proteiineissa on 2 päätyyppiä polypeptidiketjujen konformaatiota: sekundaariset ja tertiaariset rakenteet.
1. Proteiinien toissijainen rakenne
Proteiinien toissijainen rakenne- avaruudellinen rakenne, joka muodostuu peptidirungon muodostavien funktionaalisten ryhmien välisten vuorovaikutusten tuloksena. Tässä tapauksessa peptidiketjut voivat hankkia kahdentyyppisiä säännöllisiä rakenteita: α-heliksi ja β-rakenne.
?-Kierre
Tämän tyyppisessä rakenteessa peptidirunko kiertyy spiraalin muodossa johtuen vetysidosten muodostumisesta karbonyyliryhmien happiatomien ja peptidiryhmät muodostavien aminoryhmien typpiatomien välille 4 aminohappotähteen kautta. . Vetysidokset on suunnattu heliksin akselia pitkin (kuvat 1-5). On 3,6 aminohappotähdettä per turna?-heliksi.
Lähes kaikki peptidiryhmien happi- ja vetyatomit osallistuvat vetysidosten muodostukseen. Tämän seurauksena a-heliksi "supistuu" monilla vetysidoksilla. Huolimatta siitä, että nämä sidokset luokitellaan heikoiksi, niiden lukumäärä varmistaa β-heliksin maksimaalisen mahdollisen stabiilisuuden. Koska kaikki peptidirungon hydrofiiliset ryhmät osallistuvat yleensä vetysidosten muodostukseen, a-heliksien hydrofiilisyys (eli kyky muodostaa vetysidoksia veden kanssa) vähenee, kun taas niiden hydrofobisuus kasvaa.
Kierteinen rakenne on peptidirungon stabiilin konformaatio, joka vastaa vähimmäismäärää vapaata energiaa. P-heliksien muodostumisen seurauksena polypeptidiketju lyhenee, mutta jos luodaan olosuhteet vetysidosten katkeamiselle, polypeptidiketju pitenee jälleen.
Kun eri polypeptidiketjujen peptidirungon atomien välille muodostuu vetysidoksia, niitä kutsutaan juosteiden välisiksi sidoksiksi. Vetysidoksia, joita esiintyy saman polypeptidiketjun lineaaristen alueiden välillä, kutsutaan ketjunsisäisiksi. P-rakenteissa vetysidokset sijaitsevat kohtisuorassa polypeptidiketjuun nähden.
2. Proteiinien tertiäärinen rakenne
Proteiinien tertiäärinen rakenne- aminohapporadikaalien välisistä vuorovaikutuksista muodostuva kolmiulotteinen spatiaalinen rakenne, joka voi sijaita huomattavan etäisyyden päässä toisistaan polypeptidiketjussa.
Sidokset, jotka osallistuvat proteiinien tertiäärisen rakenteen muodostumiseen
Hydrofobiset vuorovaikutukset
Poimutettuna proteiinin polypeptidiketjulla on taipumus saada energeettisesti suotuisa muoto, jolle on ominaista vähimmäismäärä vapaata energiaa. Siksi aminohappojen hydrofobisilla radikaaleilla on taipumus yhdistyä vesiliukoisten proteiinien pallomaisessa rakenteessa. Niiden välissä on ns hydrofobiset vuorovaikutukset, samoin kuin van der Waalsin voimat lähellä olevien atomien välillä. Tämän seurauksena proteiinipallon sisällä hydrofobinen ydin. Peptidirungon hydrofiiliset ryhmät muodostavat sekundaarirakenteen muodostumisen aikana monia vetysidoksia, mikä sulkee pois veden sitoutumisen niihin ja proteiinin sisäisen, tiheän rakenteen tuhoutumisen.
Ioni- ja vetysidokset
Aminohappojen hydrofiilisillä radikaaleilla on taipumus muodostaa vetysidoksia veden kanssa ja siksi ne sijaitsevat pääasiassa proteiinimolekyylin pinnalla.
Kaikki hydrofobisen ytimen sisällä olevat hydrofiiliset aminohapporadikaalien ryhmät ovat vuorovaikutuksessa toistensa kanssa ioni- ja vetysidoksia käyttämällä (kuvat 1-11).
Ioniset sidokset voi esiintyä asparagiini- ja glutamiinihapporadikaalien negatiivisesti varautuneiden (anionisten) karboksyyliryhmien ja lysiini-, arginiini- tai histidiiniradikaalien positiivisesti varautuneiden (kationisten) ryhmien välillä.
Vetysidokset esiintyy hydrofiilisten varautumattomien ryhmien (kuten -OH, -CONH2, SH-ryhmien) ja minkä tahansa muiden hydrofiilisten ryhmien välillä. Ei-polaarisessa (lipidi)ympäristössä toimivilla proteiineilla, kuten kalvoproteiineilla, on käänteinen rakenne: hydrofiiliset aminohapporadikaalit sijaitsevat proteiinin sisällä, kun taas hydrofobiset aminohapot sijaitsevat molekyylin pinnalla ja ovat kosketuksissa proteiinin kanssa. ei-polaarinen ympäristö. Aminohapporadikaalit ovat kussakin tapauksessa edullisimman bioenergeettisen aseman.
kovalenttiset sidokset
Joidenkin proteiinien tertiäärinen rakenne on stabiloitunut disulfidisidokset, muodostuu kahden kysteiinitähteen SH-ryhmien vuorovaikutuksesta. Nämä kaksi kysteiinitähdettä voivat olla kaukana toisistaan proteiinin lineaarisessa primäärirakenteessa, mutta tertiäärisen rakenteen muodostuessa ne lähestyvät toisiaan ja muodostavat radikaalien vahvan kovalenttisen sidoksen.
Proteiinien kvaternäärinen rakenne
Monet proteiinit sisältävät vain yhden polypeptidiketjun. Tällaisia proteiineja kutsutaan monomeereiksi. Monomeerisiin proteiineihin kuuluvat myös proteiinit, jotka koostuvat useista ketjuista, mutta ovat liittyneet kovalenttisesti esimerkiksi disulfidisidoksilla (täten insuliinia tulee pitää monomeerisena proteiinina).
Samaan aikaan on proteiineja, jotka koostuvat kahdesta tai useammasta polypeptidiketjusta. Kunkin polypeptidiketjun kolmiulotteisen rakenteen muodostumisen jälkeen ne yhdistetään käyttämällä samoja heikkoja vuorovaikutuksia, jotka osallistuivat tertiäärisen rakenteen muodostumiseen: hydrofobinen, ioni, vety.
Polypeptidiketjujen lukumäärää ja sijaintia avaruudessa kutsutaan "proteiinien kvaternäärinen rakenne". Tällaisen proteiinin yksittäisiä polypeptidiketjuja kutsutaan protomeereiksi tai alayksiköiksi. Useita protomeerejä sisältävää proteiinia kutsutaan oligomeeriseksi.
Kaikki proteiinit, joilla on sama primäärirakenne samoissa olosuhteissa, saavat tietylle yksittäiselle proteiinille saman konformaation, joka määrittää sen spesifisen toiminnan. Proteiinin toiminnallisesti aktiivista konformaatiota kutsutaan "alkuperäinen rakenne".
klo erilaisia sairauksia kudosten proteiinikoostumuksessa tapahtuu muutos. Näitä muutoksia kutsutaan proteinopatioiksi. On olemassa perinnöllisiä ja hankittuja proteinopatioita. Perinnölliset proteinopatiat kehittyvät tietyn yksilön geneettisen laitteen vaurion seurauksena. Proteiinia ei syntetisoida ollenkaan tai se syntetisoidaan, mutta sen primäärirakenne muuttuu. Esimerkkejä perinnöllisistä proteinopatioista ovat edellä käsitellyt hemoglobinopatiat. Riippuen viallisen proteiinin roolista organismin elämässä, proteiinien konformaation ja toiminnan häiriöiden asteesta, yksilön homo- tai heterotsygoottisuudesta tälle proteiinille, perinnölliset proteinopatiat voivat aiheuttaa sairauksia, joita esiintyy vaihtelevasti. vaikeusaste, kuolemaan asti jopa ennen syntymää tai ensimmäisinä kuukausina syntymän jälkeen.
proteiinipolymorfismi - proteiinin eri muotojen olemassaolo, jotka suorittavat samoja tai hyvin samanlaisia toimintoja (isoproteiinit). Entsyymien polymorfismia (eli isoentsyymien läsnäoloa) tutkitaan useimmiten, koska ne on paljon helpompi havaita kuin muut proteiinit niiden katalysoimalla reaktiolla.
2 .Proteiinien fysikaalis-kemialliset ominaisuudet
Yksittäiset proteiinit eroavat fysikaalis-kemiallisista ominaisuuksistaan: molekyylimuodosta, molekyylipainosta, nettovarauksesta
molekyyli, natiiviproteiinimolekyylin pinnalla olevien polaaristen ja ei-polaaristen ryhmien suhde, proteiinin liukoisuus ja denaturointiaineresistenssiaste.
1. Proteiinien erot molekyylien muodossa
Kuten edellä mainittiin, molekyylien muodon mukaan proteiinit jaetaan pallomaisiin ja fibrillaarisiin. Globulaariset proteiinit ovat rakenteeltaan kompaktimpia, niiden hydrofobiset radikaalit ovat enimmäkseen piilossa hydrofobisessa ytimessä ja ne liukenevat paljon paremmin kehon nesteisiin kuin säikeiset proteiinit (poikkeuksena ovat kalvoproteiinit).
2. Proteiinien erot molekyylipainon mukaan
Proteiinit ovat korkean molekyylipainon yhdisteitä, mutta niiden molekyylipaino voi vaihdella suuresti, joka vaihtelee välillä 6 000 - 1 000 000 D ja enemmän. Proteiinin molekyylipaino riippuu polypeptidiketjussa olevien aminohappotähteiden lukumäärästä ja oligomeeristen proteiinien osalta siihen sisältyvien protomeerien (tai alayksiköiden) lukumäärästä.
3. Proteiinien kokonaisvaraus
Proteiinit sisältävät lysiini-, arginiini-, histidiini-, glutamiini- ja asparagiinihapporadikaaleja, jotka sisältävät ionisoituvia funktionaalisia ryhmiä (ionogeenisiä ryhmiä). Lisäksi polypeptidiketjujen N- ja C-päässä on a-amino- ja a-karboksyyliryhmät, jotka myös kykenevät ionisoitumaan. Proteiinimolekyylin kokonaisvaraus riippuu ionisoituneiden anionisten radikaalien Glu ja Asp sekä kationisten radikaalien Lys, Apr ja His suhteesta.
Näiden radikaalien funktionaalisten ryhmien ionisaatioaste riippuu väliaineen pH:sta. Liuoksen pH:ssa noin 7 kaikki proteiinin ionogeeniset ryhmät ovat ionisoituneessa tilassa. Happamassa ympäristössä protonien (H +) pitoisuuden kasvu johtaa karboksyyliryhmien dissosiaatioiden tukahdutukseen ja proteiinien negatiivisen varauksen vähenemiseen: -COO - + H + → -COOH. Alkalisessa ympäristössä ylimääräisen OH:n sitoutuminen protoneihin, jotka muodostuvat NH 3 +:n dissosioitumisen aikana veden muodostumisen kanssa, johtaa proteiinien positiivisen varauksen vähenemiseen:
NH3 + +OH- → -NH2 + H20.
pH-arvoa, jossa proteiini saa kokonaisnollavarauksen, kutsutaan "isoelektrinen piste" ja merkitty pI:llä. Isoelektrisessä pisteessä positiivisesti ja negatiivisesti varautuneiden proteiiniryhmien lukumäärä on sama, ts. proteiini on isoelektrisessä tilassa.
Koska useimmat solun proteiinit sisältävät enemmän anionisia ryhmiä (-COO-), näiden proteiinien isoelektrinen piste sijaitsee lievästi happamassa ympäristössä. Proteiinien isoelektrinen piste, jossa kationogeeniset ryhmät hallitsevat, on emäksisessä ympäristössä. Silmiinpistävin esimerkki tällaisista paljon arginiinia ja lysiiniä sisältävistä solunsisäisistä proteiineista ovat histonit, jotka ovat osa kromatiinia.
Proteiinit, joilla on positiivinen tai negatiivinen nettovaraus, ovat liukoisempia kuin proteiinit, jotka ovat isoelektrisessä pisteessä. Kokonaisvaraus lisää proteiinimolekyyliin sitoutuvien vesidipolien määrää ja estää samalla tavalla varautuneiden molekyylien kosketuksen, minkä seurauksena proteiinien liukoisuus paranee. Varautuneet proteiinit voivat liikkua sähkökentässä: anioniset proteiinit, joilla on negatiivinen varaus, siirtyvät kohti positiivisesti varautunutta anodia (+) ja kationiset proteiinit - kohti negatiivisesti varautunutta katodia (-). Isoelektrisessä tilassa olevat proteiinit eivät liiku sähkökentässä.
4. Polaarisen ja ei-polaarisen suhde ryhmiä luontaisten molekyylien pinnalla proteiinit
Useimpien solunsisäisten proteiinien pinnalla polaariset radikaalit hallitsevat, mutta polaaristen ja ei-polaaristen ryhmien suhde on erilainen eri proteiineille. Siten oligomeeristen proteiinien protomeerit sisältävät usein hydrofobisia radikaaleja kontaktialueella toistensa kanssa. Myös kalvojen osana toimivien tai niihin toiminnan aikana kiinnittyvien proteiinien pinnat rikastuvat hydrofobisilla radikaaleilla. Tällaiset proteiinit liukenevat paremmin lipideihin kuin veteen.
Tasoja on neljä rakenteellinen organisaatio proteiinit: primaariset, sekundaariset, tertiaariset ja kvaternaariset. Jokaisella tasolla on omat ominaisuutensa.
Proteiinien päärakenne on lineaarinen polypeptidiketju aminohapoista, jotka on yhdistetty peptidisidoksilla. Ensisijainen rakenne on proteiinimolekyylin rakenteellisen organisoinnin yksinkertaisin taso. Korkean stabiilisuuden antavat sille kovalenttiset peptidisidokset yhden aminohapon a-aminoryhmän ja toisen aminohapon a-karboksyyliryhmän välillä. [näytä] .
Jos proliinin tai hydroksiproliinin iminoryhmä on osallisena peptidisidoksen muodostumisessa, sillä on eri muoto [näytä] .

Kun soluihin muodostuu peptidisidoksia, aktivoituu ensin yhden aminohapon karboksyyliryhmä ja sitten se yhdistyy toisen aminoryhmän kanssa. Suunnilleen sama suoritetaan polypeptidien laboratorio synteesi.
Peptidisidos on polypeptidiketjun toistuva fragmentti. Sillä on useita ominaisuuksia, jotka eivät vaikuta vain primaarirakenteen muotoon, vaan myös polypeptidiketjun korkeimpiin organisoitumistasoihin:


- samantasoisuus - kaikki peptidiryhmän atomit ovat samassa tasossa;
- kyky esiintyä kahdessa resonoivassa muodossa (keto- tai enolimuoto);
- substituenttien trans-asema C-N-sidoksen suhteen;
- kyky muodostaa vetysidoksia, ja jokainen peptidiryhmä voi muodostaa kaksi vetysidosta muiden ryhmien kanssa, mukaan lukien peptidiryhmät.
Poikkeuksen muodostavat peptidiryhmät, joihin osallistuu proliinin tai hydroksiproliinin aminoryhmä. Ne pystyvät muodostamaan vain yhden vetysidoksen (katso edellä). Tämä vaikuttaa proteiinin sekundaarirakenteen muodostumiseen. Polypeptidiketju kohdassa, jossa proliini tai hydroksiproliini sijaitsee, taipuu helposti, koska sitä ei pidä, kuten tavallista, toinen vetysidos.
Peptidien ja polypeptidien nimikkeistö . Peptidien nimi muodostuu niiden aminohappojen nimistä. Kaksi aminohappoa muodostaa dipeptidin, kolme tripeptidin, neljä tetrapeptidin ja niin edelleen. Jokaisessa minkä tahansa pituisessa peptidissä tai polypeptidiketjussa on N-terminaalinen aminohappo, joka sisältää vapaan aminoryhmän, ja C-terminaalinen aminohappo, joka sisältää vapaan karboksyylin ryhmä. Polypeptidien nimeämisessä kaikki aminohapot luetellaan peräkkäin alkaen N-päästä ja korvataan niiden nimissä C-terminaalia lukuun ottamatta -in-liite -yyliin (koska peptidien aminohapoissa ei ole enää karboksyyliryhmää, mutta karbonyyli). Esimerkiksi kuvassa näkyvä nimi. 1 tripeptidi - leuc lietettä fenyylialan lietettä treoni sisään.
Proteiinin perusrakenteen ominaisuudet . Polypeptidiketjun rungossa jäykät rakenteet (litteät peptidiryhmät) vuorottelevat suhteellisen liikkuvien alueiden (-CHR) kanssa, jotka pystyvät pyörimään sidosten ympärillä. Tällaiset polypeptidiketjun rakenteen piirteet vaikuttavat sen pakkaamiseen avaruudessa.
Sekundäärinen rakenne on tapa asettaa polypeptidiketju järjestyneeksi rakenteeksi johtuen vetysidosten muodostumisesta yhden ketjun peptidiryhmien tai vierekkäisten polypeptidiketjujen välille. Konfiguraation mukaan toissijaiset rakenteet jaetaan kierteisiin (α-heliksi) ja kerroslaskostettuihin (β-rakenne ja poikki-β-muoto).


α-helix. Tämä on eräänlainen proteiinin sekundaarinen rakenne, jolla on säännöllinen kierre, joka muodostuu peptidien välisistä vetysidoksista yhdessä polypeptidiketjussa. Pauling ja Corey ehdottivat a-heliksirakennemallia (kuvio 2), joka ottaa huomioon kaikki peptidisidoksen ominaisuudet. α-heliksin pääominaisuudet:
- polypeptidiketjun kierteinen konfiguraatio, jolla on helikaalinen symmetria;
- vetysidosten muodostuminen kunkin ensimmäisen ja neljännen aminohappotähteen peptidiryhmien välille;
- spiraalin kierrosten säännöllisyys;
- kaikkien α-kierteen aminohappotähteiden vastaavuus niiden sivuradikaalien rakenteesta riippumatta;
- aminohappojen sivuradikaalit eivät osallistu α-heliksin muodostumiseen.
Ulospäin α-heliksi näyttää sähköliesi hieman venyneeltä kierteeltä. Vetysidosten säännöllisyys ensimmäisen ja neljännen peptidiryhmän välillä määrää myös polypeptidiketjun käänteiden säännöllisyyden. Yhden kierroksen korkeus tai a-heliksin nousu on 0,54 nm; se sisältää 3,6 aminohappotähdettä, eli jokainen aminohappotähde liikkuu akselia pitkin (yhden aminohappotähteen korkeus) 0,15 nm (0,54:3,6 = 0,15 nm), mikä antaa mahdollisuuden puhua kaikkien aminohappojen vastaavuudesta jäämiä α-heliksissä. α-heliksin säännöllisyysjakso on 5 kierrosta tai 18 aminohappotähdettä; yhden jakson pituus on 2,7 nm. Riisi. 3. Pauling-Coreyn α-heliksimalli
β-rakenne. Tämä on eräänlainen sekundaarinen rakenne, jolla on hieman kaareva polypeptidiketjun konfiguraatio ja joka muodostetaan käyttämällä peptidien välisiä vetysidoksia yhden polypeptidiketjun tai vierekkäisten polypeptidiketjujen erillisissä osissa. Sitä kutsutaan myös kerros-taitettu rakenteeksi. On olemassa erilaisia β-rakenteita. Proteiinin yhden polypeptidiketjun muodostamia rajoitettuja kerrosalueita kutsutaan cross-β-muodoksi (lyhyt p-rakenne). Polypeptidiketjun silmukoiden peptidiryhmien väliin muodostuu vetysidoksia ristiin-p-muodossa. Toinen tyyppi, täydellinen p-rakenne, on ominaista koko polypeptidiketjulle, jolla on pitkänomainen muoto ja jota pitävät peptidien väliset vetysidokset vierekkäisten rinnakkaisten polypeptidiketjujen välillä (kuvio 3). Tämä rakenne muistuttaa haitaripalkea. Lisäksi β-rakenteiden variantit ovat mahdollisia: ne voivat muodostua rinnakkaisista ketjuista (polypeptidiketjujen N-päät on suunnattu samaan suuntaan) ja antirinnakkaisina (N-päät on suunnattu eri suuntiin). Yhden kerroksen sivuradikaalit sijoitetaan toisen kerroksen sivuradikaalien väliin.
Proteiineissa siirtymät α-rakenteista β-rakenteisiin ja päinvastoin ovat mahdollisia vetysidosten uudelleenjärjestelyn vuoksi. Säännöllisten peptidien välisten vetysidosten sijaan ketjua pitkin (niiden takia polypeptidiketju on kierretty spiraaliksi) spiraalistetut osat ovat kierteittämättömiä ja vetysidokset suljetaan polypeptidiketjujen pitkänomaisten fragmenttien väliin. Tällainen siirtymä löytyy keratiinista, hiusproteiinista. Kun hiuksia pestään emäksisellä pesuaineella, β-keratiinin kierteinen rakenne tuhoutuu helposti ja se siirtyy α-keratiiniksi (kiharat hiukset suoristuvat).
Proteiinien säännöllisten toissijaisten rakenteiden (α-heliksit ja β-rakenteet) tuhoutumista, analogisesti kiteen sulamisen kanssa, kutsutaan polypeptidien "sulatukseksi". Tässä tapauksessa vetysidokset katkeavat ja polypeptidiketjut ovat satunnaisen kelan muodossa. Siksi sekundäärirakenteiden stabiilius määräytyvät peptidien välisillä vetysidoksilla. Muuntyyppiset sidokset eivät juuri osallistu tähän, lukuun ottamatta disulfidisidoksia polypeptidiketjussa kysteiinitähteiden kohdissa. Disulfidisidoksesta johtuvat lyhyet peptidit sulkeutuvat sykleissä. Monilla proteiineilla on samanaikaisesti α-kierteisiä alueita ja β-rakenteita. 100-prosenttisesti α-kierteestä koostuvia luonnollisia proteiineja ei juurikaan ole (poikkeus on paramyosiini, lihasproteiini, joka on 96-100-prosenttisesti α-kierre), kun taas synteettisissä polypeptideissä on 100-prosenttinen heliksi.
Muilla proteiineilla on epätasainen helicity. Paramyosiinissa, myoglobiinissa ja hemoglobiinissa havaitaan suuri taajuus a-kierteisiä rakenteita. Päinvastoin, trypsiinissä, ribonukleaasissa, merkittävä osa polypeptidiketjusta sopii kerrosrakenteisiin p-rakenteisiin. Tukikudosproteiinit: keratiinilla (hiusproteiini, villa), kollageenilla (jänneproteiini, iho), fibroiinilla (luonnollinen silkkiproteiini) on polypeptidiketjujen β-konfiguraatio. Proteiinien polypeptidiketjujen erilainen helikalisoitumisaste osoittaa, että ilmeisesti on olemassa voimia, jotka osittain häiritsevät kierteisyyttä tai "katkaisevat" polypeptidiketjun säännöllisen laskostumisen. Syynä tähän on proteiinipolypeptidiketjun tiiviimpi pakkautuminen tiettyyn tilavuuteen eli tertiääriseen rakenteeseen.
Proteiinin tertiäärinen rakenne
Proteiinin tertiäärinen rakenne on tapa, jolla polypeptidiketju laskostuu avaruudessa. Tertiäärisen rakenteen muodon mukaan proteiinit jaetaan pääasiassa pallomaisiin ja säikeisiin. Globulaarisilla proteiineilla on useimmiten elliptinen muoto, ja fibrillaariset (säikeiset) proteiinit ovat pitkänomaisia (sauvan, karan muoto).
Proteiinien tertiaarisen rakenteen konfiguraatio ei kuitenkaan vielä anna aihetta olettaa, että fibrillaarisilla proteiineilla on vain β-rakenne ja globulaarisilla α-kierteisillä. On fibrillaarisia proteiineja, joilla on kierteinen eikä kerroksittain laskostunut sekundaarinen rakenne. Esimerkiksi α-keratiini ja paramyosiini (nilviäisten sulkijalihasproteiini), tropomyosiinit (luustolihasproteiinit) ovat säikeisiä proteiineja (on sauvan muotoinen muoto) ja niiden toissijainen rakenne on a-heliksi; päinvastoin, pallomaiset proteiinit voivat sisältää suuren määrän β-rakenteita.
Lineaarisen polypeptidiketjun spiralisointi pienentää sen kokoa noin 4 kertaa; ja pakkaaminen tertiääriseen rakenteeseen tekee siitä kymmeniä kertoja kompaktimman kuin alkuperäinen ketju.
Sidokset, jotka stabiloivat proteiinin tertiaarista rakennetta . Tertiäärisen rakenteen stabiloinnissa aminohappojen sivuradikaalien välisillä sidoksilla on rooli. Nämä liitännät voidaan jakaa:
- vahva (kovalenttinen) [näytä]
.
Kovalenttisiin sidoksiin kuuluvat disulfidisidokset (-S-S-) polypeptidiketjun eri osissa sijaitsevien kysteiinien sivuradikaalien välillä; isopeptidi tai pseudopeptidi - lysiinin, arginiinin sivuradikaalien aminoryhmien, ei a-aminoryhmien, ja asparagiini-, glutamiini- ja aminositruunahappojen sivuradikaalien COOH-ryhmien välissä, ei aminohappojen a-karboksyyliryhmien välillä. Tästä johtuu tämän tyyppisen sidoksen nimi - samanlainen kuin peptidi. Harvoin löytyy esterisidosta, jonka muodostavat dikarboksyyliaminohappojen COOH-ryhmä (asparagiini, glutamiini) ja hydroksiaminohappojen OH-ryhmä (seriini, treoniini).
- heikko (polaarinen ja van der Waals) [näytä]
.
TO napasiteet sisältävät vety ja ioni. Vetysidokset syntyvät, kuten tavallista, yhden aminohapon sivuradikaalin -NH2-, -OH- tai -SH-ryhmän ja toisen aminohapon karboksyyliryhmän välillä. Ionisia tai sähköstaattisia sidoksia muodostuu sivuradikaalien -NH + 3 (lysiini, arginiini, histidiini) ja -COO - (asparagiini- ja glutamiinihappo) varautuneiden ryhmien kosketuksessa.
Ei-polaariset tai van der Waals -sidokset muodostuu aminohappojen hiilivetyradikaalien väliin. Aminohappojen alaniinin, valiinin, isoleusiinin, metioniinin ja fenyylialaniinin hydrofobiset radikaalit vuorovaikuttavat toistensa kanssa vesipitoisessa väliaineessa. Heikot van der Waalsin sidokset edistävät hydrofobisen ytimen muodostumista ei-polaarisista radikaaleista proteiinipallon sisällä. Mitä enemmän ei-polaarisia aminohappoja on, sitä suurempi on van der Waals -sidosten rooli polypeptidiketjun laskostumisessa.
Lukuisat sidokset aminohappojen sivuradikaalien välillä määräävät proteiinimolekyylin avaruudellisen konfiguraation.
Proteiinin tertiaarisen rakenteen organisoinnin piirteet . Polypeptidiketjun tertiaarisen rakenteen konformaation määräävät sen muodostavien aminohappojen sivuradikaalien ominaisuudet (joilla ei ole havaittavissa olevaa vaikutusta primääri- ja sekundaaristen rakenteiden muodostumiseen) ja mikroympäristö, eli ympäristö. Poimutettuna proteiinin polypeptidiketjulla on taipumus saada energeettisesti suotuisa muoto, jolle on ominaista vähimmäismäärä vapaata energiaa. Siksi ei-polaariset R-ryhmät, "välten" vettä, muodostavat ikään kuin proteiinin tertiaarisen rakenteen sisäosan, jossa sijaitsee pääosa polypeptidiketjun hydrofobisista tähteistä. Proteiinipallon keskellä ei ole juuri lainkaan vesimolekyylejä. Aminohapon polaariset (hydrofiiliset) R-ryhmät sijaitsevat tämän hydrofobisen ytimen ulkopuolella ja niitä ympäröivät vesimolekyylit. Polypeptidiketju taipuu omituisesti kolmiulotteisessa tilassa. Kun se taivutetaan, sekundaarinen kierteinen konformaatio katkeaa. Ketju "katkaistaan" heikoista kohdista, joissa proliini tai hydroksiproliini sijaitsevat, koska nämä aminohapot ovat liikkuvampia ketjussa muodostaen vain yhden vetysidoksen muiden peptidiryhmien kanssa. Toinen mutkan paikka on glysiini, jonka R-ryhmä on pieni (vety). Siksi muiden aminohappojen R-ryhmät pinottuna pyrkivät miehittämään vapaan tilan glysiinin sijainnissa. Useat aminohapot - alaniini, leusiini, glutamaatti, histidiini - edistävät stabiilien kierukkarakenteiden säilymistä proteiinissa, ja kuten metioniini, valiini, isoleusiini, asparagiinihappo, edistävät β-rakenteiden muodostumista. Tertiäärisen konfiguraation omaavassa proteiinimolekyylissä on osia α-heliksien (spiraalisoituneen), β-rakenteiden (kerroksisen) ja satunnaisen kierteen muodossa. Vain proteiinin oikea spatiaalinen laskostuminen tekee siitä aktiivisen; sen rikkominen johtaa proteiinin ominaisuuksien muutokseen ja biologisen aktiivisuuden menettämiseen.
Kvaternäärinen proteiinirakenne
Proteiineilla, jotka koostuvat yhdestä polypeptidiketjusta, on vain tertiäärinen rakenne. Näitä ovat myoglobiini, lihaskudoksen proteiini, joka osallistuu hapen sitomiseen, ja monet entsyymit (lysotsyymi, pepsiini, trypsiini jne.). Jotkut proteiinit on kuitenkin rakennettu useista polypeptidiketjuista, joista jokaisella on tertiäärinen rakenne. Tällaisille proteiineille on otettu käyttöön kvaternaarisen rakenteen käsite, joka tarkoittaa useiden polypeptidiketjujen, joilla on tertiäärinen rakenne, järjestäytymistä yhdeksi toiminnalliseksi proteiinimolekyyliksi. Tällaista kvaternaarisen rakenteen omaavaa proteiinia kutsutaan oligomeeriksi ja sen tertiäärisen rakenteen omaavia polypeptidiketjuja kutsutaan protomeereiksi tai alayksiköiksi (kuvio 4).

Organisaation kvaternäärisellä tasolla proteiinit säilyttävät tertiäärisen rakenteen peruskonfiguraation (pallomainen tai fibrillaarinen). Esimerkiksi hemoglobiini on proteiini, jolla on kvaternäärinen rakenne ja joka koostuu neljästä alayksiköstä. Jokainen alayksikkö on pallomainen proteiini, ja yleensä hemoglobiinilla on myös pallomainen konfiguraatio. Hiusten ja villan proteiinit - keratiinit, jotka liittyvät tertiäärisessä rakenteessa säikeisiin proteiineihin, ovat fibrillaarista ja kvaternääristä rakennetta.
Proteiinien kvaternaarisen rakenteen stabilointi . Kaikki kvaternäärisen rakenteen omaavat proteiinit eristettiin yksittäisinä makromolekyyleina, jotka eivät hajoa alayksiköiksi. Kosketukset alayksiköiden pintojen välillä ovat mahdollisia vain aminohappotähteiden polaaristen ryhmien vuoksi, koska kunkin polypeptidiketjun tertiaarisen rakenteen muodostumisen aikana ei-polaaristen aminohappojen sivuradikaalit (muodostavat suurin osa kaikista proteiinigeenisistä aminohapoista) ovat piilossa alayksikön sisällä. Polaaristen ryhmiensä väliin muodostuu lukuisia ionisia (suola-), vety- ja joissakin tapauksissa disulfidisidoksia, jotka pitävät alayksiköt lujasti organisoidun kompleksin muodossa. Vetysidoksia rikkovien tai disulfidisiltoja palauttavien aineiden käyttö aiheuttaa protomeerien hajoamista ja proteiinin kvaternaarisen rakenteen tuhoutumista. Taulukossa. Kuvassa 1 on yhteenveto tiedoista sidoksista, jotka stabiloivat proteiinimolekyylin eri organisoitumistasoja [näytä] .
| Taulukko 1. Proteiinien rakenteelliseen järjestäytymiseen osallistuvien sidosten ominaisuudet | ||
| Organisaation taso | Sidostyypit (vahvuuden mukaan) | Viestintätyyppi |
| Primaarinen (lineaarinen polypeptidiketju) | kovalenttinen (vahva) | Peptidi - aminohappojen α-amino- ja α-karboksyyliryhmien välillä |
| Toissijainen (α-heliksi, β-rakenteet) | Heikko | Vety - yhden polypeptidiketjun peptidiryhmien (jokainen ensimmäinen ja neljäs) välillä tai vierekkäisten polypeptidiketjujen peptidiryhmien välillä |
| kovalenttinen (vahva) | Disulfidi - disulfidisilmukat polypeptidiketjun lineaarisella alueella | |
| Tertiäärinen (pallomainen, fibrillaarinen) | kovalenttinen (vahva) | Disulfidi, isopeptidi, esteri - polypeptidiketjun eri osien aminohappojen sivuradikaalien välissä |
| Heikko | Vety - polypeptidiketjun eri osien aminohappojen sivuradikaalien välissä Ioni (suola) - polypeptidiketjun aminohappojen sivuradikaalien vastakkaisesti varautuneiden ryhmien välillä Van der Waals - polypeptidiketjun aminohappojen ei-polaaristen sivuradikaalien välissä |
|
| Kvaternäärinen (pallomainen, fibrillaarinen) | Heikko | Ioni - kunkin alayksikön aminohappojen sivuradikaalien vastakkaisesti varautuneiden ryhmien välillä Vety - aminohappotähteiden sivuradikaalien välillä, jotka sijaitsevat alayksiköiden kosketuksissa olevien osien pinnalla |
| kovalenttinen (vahva) | Disulfidi - eri alayksiköiden kunkin kosketuspinnan kysteiinitähteiden välillä | |
Joidenkin fibrillaaristen proteiinien rakenteellisen organisaation piirteet
Fibrillaaristen proteiinien rakenteellisella organisaatiolla on useita piirteitä verrattuna pallomaisiin proteiineihin. Nämä ominaisuudet voidaan jäljittää esimerkiksi keratiinista, fibroiinista ja kollageenista. Keratiinit esiintyvät α- ja β-konformaatioina. α-Keratiineilla ja fibroiinilla on kerroksittain laskostunut sekundaarirakenne, mutta keratiinissa ketjut ovat yhdensuuntaisia ja fibroiinissa vastasuuntaisia (katso kuva 3); lisäksi ketjujen välisiä disulfidisidoksia on keratiinissa, kun taas fibroiinissa niitä ei ole. Disulfidisidosten katkeaminen johtaa polypeptidiketjujen erottumiseen keratiineissa. Päinvastoin, maksimimäärän disulfidisidosten muodostuminen keratiineihin hapettavien aineiden vaikutuksesta luo vahvan tilarakenteen. Yleensä fibrillaarisissa proteiineissa, toisin kuin globulaarisissa proteiineissa, on joskus vaikea erottaa tiukasti toisistaan organisaation eri tasot. Jos hyväksymme (kuten globulaarisen proteiinin kohdalla), että tertiäärinen rakenne muodostuu pinoamalla yksi polypeptidiketju avaruuteen ja kvaternäärinen rakenne muodostuu useista ketjuista, niin fibrillaarisissa proteiineissa on mukana useita polypeptidiketjuja. toissijainen rakenne. Tyypillinen esimerkki fibrillaarisesta proteiinista on kollageeni, joka on yksi ihmiskehon runsaimmista proteiineista (noin 1/3 kaikkien proteiinien massasta). Sitä esiintyy kudoksissa, joissa on suuri lujuus ja heikko venyvyys (luut, jänteet, iho, hampaat jne.). Kollageenissa kolmasosa aminohappotähteistä on glysiiniä ja noin neljäsosa tai hieman enemmän proliinia tai hydroksiproliinia.

Eristetty kollageenipolypeptidiketju (primäärirakenne) näyttää katkoviivalta. Se sisältää noin 1000 aminohappoa ja sen molekyylipaino on noin 105 (kuvio 5, a, b). Polypeptidiketju rakennetaan toistuvasta aminohappotripletistä (tripletti), jonka koostumus on seuraava: gly-A-B, jossa A ja B ovat mitä tahansa aminohappoja paitsi glysiiniä (useimmiten proliinia ja hydroksiproliinia). Kollageenipolypeptidiketjut (tai a-ketjut) sekundaari- ja tertiääristen rakenteiden muodostumisen aikana (kuvio 5, c ja d) eivät voi muodostaa tyypillisiä a-heliksejä, joilla on kierteinen symmetria. Tämän estävät proliini, hydroksiproliini ja glysiini (antikierteiset aminohapot). Siksi kolme α-ketjua muodostavat ikään kuin kierrettyjä spiraaleja, kuten kolme lankaa, jotka kiertyvät sylinterin ympärille. Kolme kierteistä α-ketjua muodostavat toistuvan kollageenirakenteen, jota kutsutaan tropokollageeniksi (kuva 5d). Tropokollageeni on organisaatiossaan kollageenin tertiäärinen rakenne. Proliinin ja hydroksiproliinin litteät renkaat, jotka vaihtelevat säännöllisesti pitkin ketjua, antavat sille jäykkyyttä, samoin kuin ketjujen väliset sidokset tropokollageenin α-ketjujen välillä (siksi kollageeni kestää venymistä). Tropokollageeni on pohjimmiltaan kollageenifibrillien alayksikkö. Tropokollageenialayksiköt pinoutuvat kollageenin kvaternaariseen rakenteeseen vaiheittain (kuvio 5e).
Kollageenirakenteiden stabiloituminen tapahtuu ketjujen välisten vety-, ioni- ja van der Waalsin sidosten sekä pienen määrän kovalenttisten sidosten ansiosta.
Kollageenin α-ketjuilla on erilaiset kemialliset rakenteet. Erottele α 1 -ketjut eri tyyppejä(I, II, III, IV) ja a2-ketjut. Riippuen siitä, mitkä α 1 - ja α 2 -ketjut osallistuvat tropokollageenin kolmijuosteisen heliksin muodostumiseen, kollageenia erotetaan neljää tyyppiä:
- ensimmäinen tyyppi - kaksi a1 (I) ja yksi a2-ketju;
- toinen tyyppi - kolme a1(II)-ketjua;
- kolmas tyyppi - kolme a1(III)-ketjua;
- neljäs tyyppi - kolme a1(IV)-ketjua.
Yleisin ensimmäisen tyypin kollageeni: sitä löytyy luukudoksesta, ihosta, jänteistä; tyypin 2 kollageenia löytyy rustosta jne. Samassa kudoksessa voi olla erilaisia kollageenityyppejä.
Kollageenirakenteiden järjestynyt aggregaatio, niiden jäykkyys ja inertisyys tarjoavat kollageenikuitujen korkean lujuuden. Kollageeniproteiinit sisältävät myös hiilihydraattikomponentteja, eli ne ovat proteiini-hiilihydraattikomplekseja.
Kollageeni on solunulkoinen proteiini, jonka muodostavat sidekudoksen solut, jotka ovat osa kaikkia elimiä. Siksi kollageenin vaurioituessa (tai sen muodostumisen rikkoutuessa) esiintyy useita elinten sidekudoksen tukitoimintojen rikkomuksia.
| Sivu 3 | sivuja yhteensä: 7 |
MODUULI 1 PROTEIINIEN RAKENNE, OMINAISUUDET JA TOIMINNOT
MODUULI 1 PROTEIINIEN RAKENNE, OMINAISUUDET JA TOIMINNOT
Moduulin rakenne | Teemat |
Modulaarinen yksikkö 1 | 1.1. Proteiinien rakenneorganisaatio. Proteiinien luonnollisen konformaation muodostumisvaiheet 1.2. Proteiinin toiminnan perusteet. Lääkkeet ligandeina, jotka vaikuttavat proteiinien toimintaan 1.3. Proteiinien denaturaatio ja niiden spontaanin renaturoitumisen mahdollisuus |
Modulaarinen yksikkö 2 | 1.4. Oligomeeristen proteiinien rakenteen ja toiminnan piirteet hemoglobiinin esimerkissä 1.5. Proteiinien luonnollisen konformaation ylläpitäminen solussa 1.6. Erilaisia proteiineja. Proteiiniperheet immunoglobuliinien esimerkissä 1.7. Proteiinien fysikaalis-kemialliset ominaisuudet ja menetelmät niiden erottamiseksi |
Moduuliyksikkö 1 MONOMEERISTEN PROTEIINIEN RAKENNEJÄRJESTELY JA NIIDEN TOIMINNAN PERUSTA
Oppimistavoitteet Kykyä:
1. Käytä tietoa proteiinien rakenteellisista ominaisuuksista ja proteiinien toimintojen riippuvuudesta niiden rakenteesta ymmärtääksesi perinnöllisten ja hankittujen proteinopatioiden kehittymismekanismeja.
2. Selitä tiettyjen lääkkeiden terapeuttisen vaikutuksen mekanismeja ligandeina, jotka ovat vuorovaikutuksessa proteiinien kanssa ja muuttavat niiden aktiivisuutta.
3. Käytä tietoa proteiinien rakenteesta ja konformationaalisesta labilisuudesta ymmärtääksesi niiden rakenteellista ja toiminnallista epästabiilisuutta ja taipumusta denaturoitua muuttuvissa olosuhteissa.
4. Selitä denaturointiaineiden käyttö lääketieteellisten materiaalien ja instrumenttien steriloinnissa sekä antiseptisinä aineina.
Tietää:
1. Proteiinien rakenteellisen organisoinnin tasot.
2. Proteiinien primäärirakenteen merkitys, joka määrää niiden rakenteellisen ja toiminnallisen monimuotoisuuden.
3. Proteiinien aktiivisen keskuksen muodostumismekanismi ja sen spesifinen vuorovaikutus ligandin kanssa, joka on proteiinien toiminnan taustalla.
4. Esimerkkejä eksogeenisten ligandien (lääkkeet, toksiinit, myrkyt) vaikutuksesta proteiinien konformaatioon ja toiminnalliseen aktiivisuuteen.
5. Proteiinien denaturoitumisen syyt ja vaikutukset, denaturaatiota aiheuttavat tekijät.
6. Esimerkkejä denaturoivien tekijöiden käytöstä lääketieteessä antiseptisinä aineina ja keinoina lääketieteellisten instrumenttien sterilointiin.
AIHE 1.1. PROTEIINIEN RAKENNEJÄRJESTELY. ALKUPERÄISEN MUODOSTAMINEN
PROTEIINIKONFORMAATIOT
Proteiinit ovat polymeerimolekyylejä, joiden monomeerit ovat vain 20 a-aminohappoa. Proteiinissa olevien aminohappojen sarjan ja liittymisjärjestyksen määrää yksilöiden DNA:ssa olevien geenien rakenne. Jokainen proteiini suorittaa oman tehtävänsä erityisrakenteensa mukaisesti. Tietyn organismin proteiinien joukko määrittää sen fenotyyppiset ominaisuudet, samoin kuin perinnöllisten sairauksien esiintymisen tai taipumuksen niiden kehittymiseen.
1. Aminohapot, jotka muodostavat proteiineja. peptidisidos. Proteiinit ovat polymeerejä, jotka on rakennettu monomeereistä - 20 α-aminohaposta, joiden yleinen kaava on
Aminohapot eroavat α-hiiliatomiin kiinnittyneiden radikaalien rakenteesta, koosta ja fysikaalis-kemiallisista ominaisuuksista. Aminohappojen funktionaaliset ryhmät määrittävät eri α-aminohappojen ominaisuuksien piirteet. α-aminohapoissa esiintyvät radikaalit voidaan jakaa useisiin ryhmiin:
proliini, toisin kuin muut 19 proteiinimonomeeriä, ei aminohappo, vaan iminohappo, proliinissa oleva radikaali liittyy sekä α-hiiliatomiin että iminoryhmään
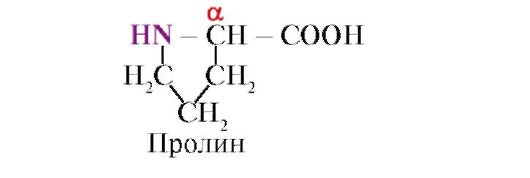 Aminohapot eroavat toisistaan liukoisuudessaan veteen. Tämä johtuu radikaalien kyvystä olla vuorovaikutuksessa veden kanssa (hydratoitua).
Aminohapot eroavat toisistaan liukoisuudessaan veteen. Tämä johtuu radikaalien kyvystä olla vuorovaikutuksessa veden kanssa (hydratoitua).
TO hydrofiilinen Käsitteet sisältävät anionisia, kationisia ja polaarisia varautumattomia funktionaalisia ryhmiä sisältävät radikaalit.
TO hydrofobinen Käsitteet sisältävät radikaaleja, jotka sisältävät metyyliryhmiä, alifaattisia ketjuja tai syklejä.
2. Peptidisidokset yhdistävät aminohapot peptideiksi. Peptidin synteesin aikana yhden aminohapon α-karboksyyliryhmä on vuorovaikutuksessa toisen aminohapon α-aminoryhmän kanssa muodostaen peptidisidos:
Proteiinit ovat polypeptidejä, ts. α-aminohappojen lineaariset polymeerit, jotka on yhdistetty peptidisidoksella (kuva 1.1.)
 Riisi. 1.1. Termit, joita käytetään kuvaamaan peptidien rakennetta
Riisi. 1.1. Termit, joita käytetään kuvaamaan peptidien rakennetta
Aminohappomonomeerejä, jotka muodostavat polypeptidejä, kutsutaan aminohappotähteet. Toistuvien ryhmien ketju - NH-CH-CO- lomakkeet peptidirunko. Aminohappotähdettä, jossa on vapaa a-aminoryhmä, kutsutaan N-päätteeksi, ja aminohappotähdettä, jossa on vapaa a-karboksyyliryhmä, kutsutaan C-päätteeksi. Peptidit kirjoitetaan ja luetaan N-päästä C-päähän.
Proliinin iminoryhmän muodostama peptidisidos eroaa muista peptidisidoksista: peptidiryhmän typpiatomista puuttuu vety,
sen sijaan on olemassa sidos radikaalin kanssa, minkä seurauksena syklin toinen puoli sisältyy peptidirunkoon:
 Peptidit eroavat toisistaan aminohappokoostumuksessa, aminohappojen lukumäärässä ja aminohappojen järjestyksessä, esimerkiksi Ser-Ala-Glu-Gis ja His-Glu-Ala-Ser ovat kaksi eri peptidiä.
Peptidit eroavat toisistaan aminohappokoostumuksessa, aminohappojen lukumäärässä ja aminohappojen järjestyksessä, esimerkiksi Ser-Ala-Glu-Gis ja His-Glu-Ala-Ser ovat kaksi eri peptidiä.
Peptidisidokset ovat erittäin vahvoja, ja niiden kemiallinen ei-entsymaattinen hydrolyysi vaatii ankaria olosuhteita: analysoitua proteiinia hydrolysoidaan väkevässä suolahapossa noin 110°C:n lämpötilassa 24 tunnin ajan. Elävässä solussa peptidisidokset voivat katketa proteolyyttiset entsyymit, nimeltään proteaasit tai peptidihydrolaasit.
3. Proteiinien perusrakenne. Aminohappotähteet eri proteiinien peptidiketjuissa eivät vuorottele satunnaisesti, vaan ne on järjestetty tiettyyn järjestykseen. Lineaarista sekvenssiä tai aminohappotähteiden sekvenssiä polypeptidiketjussa kutsutaan proteiinin ensisijainen rakenne.
Kunkin yksittäisen proteiinin primäärirakenne on koodattu DNA-molekyylissä (alueella, jota kutsutaan geeniksi) ja se toteutuu transkription (mRNA:n tietojen uudelleenkirjoittaminen) ja translaation (proteiinin primäärirakenteen synteesi) aikana. Näin ollen yksittäisen ihmisen proteiinien primäärirakenne on vanhemmilta lapsille peritty tieto, joka määrää tietyn organismin proteiinien rakenteelliset ominaisuudet, joista olemassa olevien proteiinien toiminta riippuu (kuva 1.2.).
 Riisi. 1.2. Genotyypin ja yksilön kehossa syntetisoitujen proteiinien konformaation välinen suhde
Riisi. 1.2. Genotyypin ja yksilön kehossa syntetisoitujen proteiinien konformaation välinen suhde
Jokaisella ihmiskehon noin 100 000 yksittäisestä proteiinista on ainutlaatuinen ensisijainen rakenne. Yhden tyyppisen proteiinin (esimerkiksi albumiinin) molekyyleissä on sama aminohappotähteiden vuorottelu, mikä erottaa albumiinin kaikista muista yksittäisistä proteiineista.
Peptidiketjun aminohappotähteiden sekvenssiä voidaan pitää eräänä tiedontallennusmuotona. Tämä tieto määrää lineaarisen peptidiketjun avaruudellisen laskostumisen kompaktimmaksi kolmiulotteiseksi rakenteeksi ns. konformaatio orava. Funktionaalisesti aktiivisen proteiinikonformaation muodostumisprosessia kutsutaan taitettava.
4. Proteiinien konformaatio. Vapaa pyöriminen peptidirungossa on mahdollista peptidiryhmän typpiatomin ja viereisen a-hiiliatomin sekä a-hiiliatomin ja karbonyyliryhmän hiilen välillä. Aminohappotähteiden funktionaalisten ryhmien vuorovaikutuksen ansiosta proteiinien primäärirakenne voi saada monimutkaisempia tilarakenteita. Globulaarisissa proteiineissa peptidiketjujen konformaation laskostumisen kaksi päätasoa erotetaan: toissijainen Ja tertiäärinen rakenne.
Proteiinien toissijainen rakenne- tämä on avaruudellinen rakenne, joka muodostuu vetysidosten muodostumisen seurauksena peptidirungon funktionaalisten ryhmien -C=O ja -NH- välille. Tässä tapauksessa peptidiketju voi hankkia kahdentyyppisiä säännöllisiä rakenteita: α-heliksit Ja β rakenteet.
SISÄÄN α-heliksit vetysidokset muodostuvat karbonyyliryhmän happiatomin ja siitä peräisin olevan 4. aminohapon amiditypen vedyn välille; aminohappotähteiden sivuketjut
sijaitsee kierteen reunalla, ei osallistu sekundaarirakenteen muodostukseen (kuva 1.3.).
Kookkaat radikaalit tai radikaalit, joissa on samat varaukset, estävät a-heliksin muodostumisen. Proliinitähde, jolla on rengasrakenne, katkaisee a-heliksin, koska vedyn puuttumisen vuoksi peptidiketjun typpiatomissa on mahdotonta muodostaa vetysidosta. Typen ja a-hiiliatomin välinen sidos on osa proliinisykliä, joten peptidirunko saa mutkan tässä paikassa.
β-rakenne muodostuu yhden polypeptidiketjun peptidirungon lineaaristen alueiden väliin, jolloin muodostuu laskostettuja rakenteita. Polypeptidiketjuja tai niiden osia voi muodostua rinnakkain tai antirinnakkaiset β-rakenteet. Ensimmäisessä tapauksessa vuorovaikutuksessa olevien peptidiketjujen N- ja C-päät ovat yhtenevät, ja toisessa tapauksessa niillä on päinvastainen suunta (kuva 1.4).
Riisi. 1.3. Proteiinin sekundaarirakenne - α-heliksi
 Riisi. 1.4. Rinnakkaiset ja vastasuuntaiset β-laskostetut rakenteet
Riisi. 1.4. Rinnakkaiset ja vastasuuntaiset β-laskostetut rakenteet
β-rakenteet on merkitty leveillä nuolilla: A - Vastasuuntainen β-rakenne. B - Yhdensuuntaiset β-laskostetut rakenteet
Joissakin proteiineissa β-rakenteita voi muodostua johtuen vetysidosten muodostumisesta eri polypeptidiketjujen peptidirungon atomien välille.
Löytyy myös proteiineista alueet, joilla on epäsäännöllinen toissijainen rakenne, joka sisältää polypeptidirungon mutkia, silmukoita ja käänteitä. Ne sijaitsevat usein paikoissa, joissa peptidiketjun suunta muuttuu, esimerkiksi rinnakkaisen β-levyrakenteen muodostumisen aikana.
α-heliksien ja β-rakenteiden läsnäolon perusteella pallomaiset proteiinit voidaan jakaa neljään luokkaan.
Riisi. 1.5. Myoglobiinin (A) ja hemoglobiinin β-ketjun (B) toissijainen rakenne, joka sisältää kahdeksan α-heliksiä

 Riisi. 1.6. Trioosifosfaatti-isomeraasin ja pyruvaattikinaasidomeenin toissijainen rakenne
Riisi. 1.6. Trioosifosfaatti-isomeraasin ja pyruvaattikinaasidomeenin toissijainen rakenne
 Riisi. 1.7. Immunoglobuliinin vakiodomeenin (A) ja superoksididismutaasientsyymin (B) toissijainen rakenne
Riisi. 1.7. Immunoglobuliinin vakiodomeenin (A) ja superoksididismutaasientsyymin (B) toissijainen rakenne
SISÄÄN neljäs luokka sisälsi proteiineja, joiden koostumuksessa on pieni määrä säännöllisiä sekundaarisia rakenteita. Nämä proteiinit sisältävät pieniä, kysteiinipitoisia proteiineja tai metalloproteiineja.
Proteiinin tertiäärinen rakenne- aminohapporadikaalien välisistä vuorovaikutuksista muodostuva konformaatiotyyppi, joka voi sijaita huomattavan etäisyyden päässä toisistaan peptidiketjussa. Tässä tapauksessa useimmat proteiinit muodostavat spatiaalisen rakenteen, joka muistuttaa palloa (globulaariset proteiinit).
Koska aminohappojen hydrofobisilla radikaaleilla on taipumus yhdistyä ns hydrofobisia vuorovaikutuksia ja molekyylien välisten van der Waalsin voimien vuoksi proteiinipallon sisään muodostuu tiheä hydrofobinen ydin. Hydrofiiliset ionisoidut ja ionisoimattomat radikaalit sijaitsevat pääasiassa proteiinin pinnalla ja määrittävät sen vesiliukoisuuden.
 Riisi. 1.8 Sidostyypit, jotka syntyvät aminohapporadikaalien välille proteiinin tertiaarisen rakenteen muodostumisen aikana
Riisi. 1.8 Sidostyypit, jotka syntyvät aminohapporadikaalien välille proteiinin tertiaarisen rakenteen muodostumisen aikana
1 - ionisidos- esiintyy positiivisesti ja negatiivisesti varautuneiden funktionaalisten ryhmien välillä;
2 - vetysidos- esiintyy hydrofiilisen varautumattoman ja minkä tahansa muun hydrofiilisen ryhmän välillä;
3 - hydrofobisia vuorovaikutuksia- esiintyä hydrofobisten radikaalien välillä;
4 - disulfidisidos- muodostuu kysteiinitähteiden SH-ryhmien hapettumisesta ja niiden vuorovaikutuksesta keskenään
Hydrofiiliset aminohappotähteet hydrofobisen ytimen sisällä voivat olla vuorovaikutuksessa toistensa kanssa käyttämällä ioninen Ja vetysidoksia(Kuva 1.8).
Ioni- ja vetysidokset sekä hydrofobiset vuorovaikutukset ovat heikkoja: niiden energia ylittää hieman molekyylien lämpöliikkeen energian huoneenlämpötilassa. Proteiinin konformaatiota ylläpitää monien tällaisten heikkojen sidosten esiintyminen. Koska proteiinin muodostavat atomit ovat jatkuvassa liikkeessä, on mahdollista rikkoa joitakin heikkoja sidoksia ja muodostaa muita, mikä johtaa polypeptidiketjun yksittäisten osien pieniin liikkeisiin. Tätä proteiinien ominaisuutta muuttaa konformaatiota joidenkin hajoamisen ja muiden heikkojen sidosten muodostumisen seurauksena kutsutaan konformationaalinen labilisuus.
Ihmiskehossa on järjestelmät, jotka tukevat homeostaasi- pysyvyys sisäinen ympäristö tietyissä terveen kehon hyväksyttävissä rajoissa. Homeostaasin olosuhteissa pienet muutokset konformaatiossa eivät häiritse proteiinien yleistä rakennetta ja toimintaa. Proteiinin toiminnallisesti aktiivista konformaatiota kutsutaan syntyperäinen rakenne. Muutos sisäisessä ympäristössä (esim. glukoosin, Ca-ionien, protonien jne. pitoisuus) johtaa proteiinien konformaation muutokseen ja toiminnan häiriintymiseen.
Joidenkin proteiinien tertiäärinen rakenne on stabiloitunut disulfidisidokset, muodostuu kahden tähteen -SH-ryhmien vuorovaikutuksesta
 Riisi. 1.9. Disulfidisidoksen muodostuminen proteiinimolekyylissä
Riisi. 1.9. Disulfidisidoksen muodostuminen proteiinimolekyylissä
kysteiini (kuva 1.9). Useimpien solunsisäisten proteiinien tertiäärisessä rakenteessa ei ole kovalenttisia disulfidisidoksia. Niiden läsnäolo on ominaista solun erittämille proteiineille, mikä varmistaa niiden paremman stabiilisuuden solunulkoisissa olosuhteissa. Joten disulfidisidoksia on läsnä insuliinin ja immunoglobuliinien molekyyleissä.
Insuliini- proteiinihormoni, joka syntetisoituu haiman β-soluissa ja erittyy vereen vasteena veren glukoosipitoisuuden nousulle. Insuliinin rakenteessa on kaksi disulfidisidosta, jotka yhdistävät polypeptidin A- ja B-ketjut, ja yksi disulfidisidos A-ketjun sisällä (kuva 1.10).
 Riisi. 1.10. Disulfidisidokset insuliinin rakenteessa
Riisi. 1.10. Disulfidisidokset insuliinin rakenteessa
5. Proteiinien supersekundaarinen rakenne. Proteiineissa, jotka ovat toisinaan erilaisia primäärirakenteeltaan ja -toiminnoiltaan samanlaiset yhdistelmät ja sekundäärirakenteiden sijoittaminen toisiinsa, joita kutsutaan ylisekundaariseksi rakenteeksi. Se on väliasemassa sekundääristen ja tertiääristen rakenteiden välillä, koska se on erityinen yhdistelmä sekundäärisiä rakenne-elementtejä proteiinin tertiaarisen rakenteen muodostumisen aikana. Supersekundaarisilla rakenteilla on erityiset nimet, kuten "a-helix-turn-a-helix", "leusiinivetoketju", "sinkkisormet" jne. Tällaiset supersekundaariset rakenteet ovat ominaisia DNA:ta sitoville proteiineille.
"Leusiinivetoketju". Tällaista supersekundaarista rakennetta käytetään yhdistämään kaksi proteiinia. Vuorovaikutteisten proteiinien pinnalla on α-kierteisiä alueita, jotka sisältävät vähintään neljä leusiinitähdettä. Leusiinitähteet α-heliksissä sijaitsevat kuuden aminohapon päässä toisistaan. Koska jokainen α-heliksin kierros sisältää 3,6 aminohappotähdettä, leusiiniradikaaleja löytyy joka toisen kierroksen pinnasta. Yhden proteiinin α-heliksin leusiinitähteet voivat olla vuorovaikutuksessa toisen proteiinin leusiinitähteiden kanssa (hydrofobiset vuorovaikutukset), jotka yhdistävät ne toisiinsa (kuva 1.11.). Monet DNA:ta sitovat proteiinit toimivat osana oligomeerikomplekseja, joissa yksittäiset alayksiköt on liitetty toisiinsa "leusiinivetoketjuilla".
 Riisi. 1.11. "Leusiinivetoketju" kahden proteiinin α-kierteisten alueiden välissä
Riisi. 1.11. "Leusiinivetoketju" kahden proteiinin α-kierteisten alueiden välissä
Histonit ovat esimerkki tällaisista proteiineista. Histonit- ydinproteiinit, jotka sisältävät suuren määrän positiivisesti varautuneita aminohappoja - arginiini ja lysiini (jopa 80%). Histonimolekyylit yhdistetään oligomeerisiksi komplekseiksi, jotka sisältävät kahdeksan monomeeriä "leusiinikiinnittimien" avulla huolimatta näiden molekyylien merkittävästä homonyymistä varauksesta.
"sinkkisormi"- DNA:ta sitoville proteiineille tyypillinen supersekundaarirakenteen variantti on proteiinin pinnalla pitkänomaisen fragmentin muotoinen ja sisältää noin 20 aminohappotähdettä (kuva 1.12). "Vennytetyn sormen" muotoa tukee sinkkiatomi, joka liittyy neljään aminohapporadikaaliin - kahteen kysteiinitähteeseen ja kahteen histidiinitähteeseen. Joissakin tapauksissa histidiinitähteiden sijasta on kysteiinijäämiä. Kaksi lähekkäin olevaa kysteiinitähdettä erotetaan kahdesta muusta Gisili-tähteestä noin 12 aminohappotähteen Cys-sekvenssillä. Tämä proteiinin alue muodostaa a-heliksin, jonka radikaalit voivat sitoutua spesifisesti DNA:n pääuran säätelyalueisiin. Yksilön sitoutumisen spesifisyys
 Riisi. 1.12. DNA:ta sitovien proteiinien osan ensisijainen rakenne, joka muodostaa "sinkkisormen" rakenteen (kirjaimet osoittavat aminohappoja, jotka muodostavat tämän rakenteen)
Riisi. 1.12. DNA:ta sitovien proteiinien osan ensisijainen rakenne, joka muodostaa "sinkkisormen" rakenteen (kirjaimet osoittavat aminohappoja, jotka muodostavat tämän rakenteen)
säätelevä DNA:ta sitova proteiini riippuu "sinkkisormessa" sijaitsevien aminohappotähteiden sekvenssistä. Tällaiset rakenteet sisältävät erityisesti steroidihormonireseptoreita, jotka osallistuvat transkription säätelyyn (informaation lukeminen DNA:sta RNA:ksi).
AIHE 1.2. PROTEIINITOIMINNAN PERUSTEET. LAIMET LIGANDEINA, JOTKA VAIKUTTAVAT PROTEIIINITOIMINTOA
1. Proteiinin aktiivinen keskus ja sen vuorovaikutus ligandin kanssa. Tertiäärisen rakenteen muodostumisen aikana funktionaalisesti aktiivisen proteiinin pinnalle, yleensä syvennykseen, muodostuu paikka primäärirakenteessa kaukana toisistaan olevista aminohapporadikaaleista. Tätä kohtaa, jolla on ainutlaatuinen rakenne tietylle proteiinille ja joka pystyy spesifisesti vuorovaikuttamaan tietyn molekyylin tai samankaltaisten molekyylien ryhmän kanssa, kutsutaan proteiinin sitomispaikaksi ligandin tai aktiivisen kohdan kanssa. Ligandit ovat molekyylejä, jotka ovat vuorovaikutuksessa proteiinien kanssa.
Korkea spesifisyys Proteiinin vuorovaikutus ligandin kanssa varmistetaan aktiivisen keskuksen rakenteen komplementaarisella ligandin rakenteen kanssa.
täydentävyyttä on vuorovaikutuksessa olevien pintojen avaruudellinen ja kemiallinen vastaavuus. Aktiivisen keskuksen tulee paitsi avaruudellisesti vastata siihen kuuluvaa ligandia, myös aktiiviseen keskukseen sisältyvien radikaalien funktionaalisten ryhmien ja ligandin välille tulee muodostua sidoksia (ioni-, vety- ja hydrofobiset vuorovaikutukset), jotka pitävät ligandin. aktiivisessa keskustassa (kuva 1.13 ).
 Riisi. 1.13. Proteiinin täydentävä vuorovaikutus ligandin kanssa
Riisi. 1.13. Proteiinin täydentävä vuorovaikutus ligandin kanssa
Joillakin ligandeilla, kun ne ovat kiinnittyneet proteiinin aktiiviseen keskukseen, on apurooli proteiinien toiminnassa. Tällaisia ligandeja kutsutaan kofaktoreiksi, ja proteiineja, joiden koostumuksessa on ei-proteiininen osa, kutsutaan monimutkaiset proteiinit(toisin kuin yksinkertaiset proteiinit, jotka koostuvat vain proteiiniosasta). Proteiiniin lujasti kiinnittyvää ei-proteiiniosaa kutsutaan prosteettinen ryhmä. Esimerkiksi myoglobiinin, hemoglobiinin ja sytokromien koostumus sisältää proteettisen ryhmän, joka on kiinteästi kiinnittynyt aktiiviseen keskustaan - rauta-ionin sisältävän hemin. Hemeä sisältäviä monimutkaisia proteiineja kutsutaan hemoproteiineiksi.
Kun spesifisiä ligandeja kiinnittyy proteiineihin, näiden proteiinien toiminta ilmenee. Siten albumiini, veriplasman tärkein proteiini, osoittaa kuljetustoimintonsa kiinnittämällä aktiiviseen keskukseen hydrofobisia ligandeja, kuten rasvahappoja, bilirubiinia, joitain lääkkeitä jne. (Kuva 1.14)
Peptidiketjun kolmiulotteisen rakenteen kanssa vuorovaikutuksessa olevat ligandit voivat olla paitsi pienimolekyylipainoisia orgaanisia ja epäorgaanisia molekyylejä, myös makromolekyylejä:
DNA (yllä käsitellyt esimerkit DNA:ta sitovien proteiinien kanssa);
polysakkaridit;
 Riisi. 1.14. Genotyypin ja fenotyypin välinen suhde
Riisi. 1.14. Genotyypin ja fenotyypin välinen suhde
DNA-molekyyliin koodattu ihmisen proteiinien ainutlaatuinen primäärirakenne toteutuu soluissa ainutlaatuisen konformaation, aktiivisen kohdan rakenteen ja proteiinitoimintojen muodossa.
Näissä tapauksissa proteiini tunnistaa ligandin spesifisen alueen, joka on suhteessa sitoutumiskohtaan ja täydentää sitä. Hepatosyyttien pinnalla on siis insuliinihormonin reseptoriproteiineja, jolla on myös proteiinirakenne. Insuliinin vuorovaikutus reseptorin kanssa aiheuttaa muutoksen sen konformaatiossa ja signaalijärjestelmien aktivoitumisen, mikä johtaa ravinteiden kertymiseen maksasoluihin syömisen jälkeen.
Täten, Proteiinien toiminta perustuu proteiinin aktiivisen keskuksen spesifiseen vuorovaikutukseen ligandin kanssa.
2. Domeenirakenne ja sen rooli proteiinien toiminnassa. Globulaaristen proteiinien pitkät polypeptidiketjut laskostuvat usein useiksi kompakteiksi, suhteellisen itsenäisiksi alueiksi. Niillä on itsenäinen tertiäärinen rakenne, joka muistuttaa globulaaristen proteiinien rakennetta, ja niitä kutsutaan verkkotunnuksia. Proteiinien domeenirakenteen ansiosta niiden tertiäärinen rakenne on helpompi muodostaa.
Domeeniproteiineissa ligandia sitovat kohdat sijaitsevat usein domeenien välissä. Joten trypsiini on proteolyyttinen entsyymi, jota haiman eksokriininen osa tuottaa ja joka on välttämätön ruokaproteiinien sulattamiseksi. Sillä on kaksidomeenirakenne, ja trypsiinin sitoutumiskohta ligandinsa - ruokaproteiinin - kanssa sijaitsee näiden kahden domeenin välisessä urassa. Aktiivisessa keskustassa luodaan olosuhteet, jotka ovat välttämättömiä elintarvikeproteiinin tietyn kohdan tehokkaalle sitoutumiselle ja sen peptidisidosten hydrolyysille.
Proteiinin eri domeenit voivat liikkua suhteessa toisiinsa, kun aktiivinen keskus on vuorovaikutuksessa ligandin kanssa (kuva 1.15).
Heksokinaasi- entsyymi, joka katalysoi glukoosin fosforylaatiota ATP:n avulla. Entsyymin aktiivinen kohta sijaitsee kahden domeenin välisessä rakossa. Kun heksokinaasi sitoutuu glukoosiin, ympäröivät domeenit sulkeutuvat ja substraatti jää loukkuun, jossa tapahtuu fosforylaatiota (katso kuva 1.15).
 Riisi. 1.15. Heksokinaasidomeenien sitoutuminen glukoosiin
Riisi. 1.15. Heksokinaasidomeenien sitoutuminen glukoosiin
Joissakin proteiineissa domeenit suorittavat itsenäisiä toimintoja sitoutumalla erilaisiin ligandeihin. Tällaisia proteiineja kutsutaan monitoimisiksi.
3. Lääkkeet - ligandit, jotka vaikuttavat proteiinien toimintaan. Proteiinien vuorovaikutus ligandien kanssa on spesifistä. Proteiinin ja sen aktiivisen kohdan konformaatiolabiteetin vuoksi on kuitenkin mahdollista valita toinen aine, joka voisi myös olla vuorovaikutuksessa aktiivisessa kohdassa tai molekyylin toisessa osassa olevan proteiinin kanssa.
Ainetta, joka on rakenteeltaan samanlainen kuin luonnollinen ligandi, kutsutaan ligandin rakenteellinen analogi tai luonnoton ligandi. Se on myös vuorovaikutuksessa aktiivisessa kohdassa olevan proteiinin kanssa. Ligandin rakenteellinen analogi voi molemmat parantaa proteiinin toimintaa (agonisti) ja vähentää sitä (antagonisti). Ligandi ja sen rakenteelliset analogit kilpailevat toistensa kanssa proteiinin sitoutumisesta samassa kohdassa. Tällaisia aineita kutsutaan kilpailukykyiset modulaattorit proteiinitoimintojen säätelijät. monet lääkkeitä toimivat proteiinin estäjinä. Jotkut niistä saadaan luonnollisten ligandien kemiallisella modifioinnilla. Proteiinitoiminnan estäjät voivat olla lääkkeitä ja myrkkyjä.
Atropiini on M-kolinergisten reseptorien kilpaileva estäjä. Asetyylikoliini on välittäjäaine hermoimpulssien välittämiseen kolinergisten synapsien kautta. Virityksen suorittamiseksi synaptiseen rakoon vapautuvan asetyylikoliinin on oltava vuorovaikutuksessa proteiinin - postsynaptisen kalvon reseptorin - kanssa. Kaksi tyyppiä löytyi kolinergiset reseptorit:
M-reseptori asetyylikoliinin lisäksi se vuorovaikuttaa selektiivisesti muskariinin (kärpäshelttatoksiinin) kanssa. M - kolinergiset reseptorit ovat läsnä sileissä lihaksissa ja vuorovaikutuksessa asetyylikoliinin kanssa aiheuttavat niiden supistumisen;
H-reseptori sitoutuu erityisesti nikotiiniin. N-kolinergisiä reseptoreita löytyy poikkijuovaisten luustolihasten synapseista.
spesifinen estäjä M-kolinergiset reseptorit on atropiini. Sitä löytyy belladonna- ja henbane-kasveista.
 Atropiinissa on rakenteeltaan asetyylikoliinin kaltaisia funktionaalisia ryhmiä ja niiden tilajärjestelyjä, joten se kuuluu M-kolinergisten reseptorien kilpaileviin estäjiin. Koska asetyylikoliinin sitoutuminen M-kolinergisiin reseptoreihin aiheuttaa sileiden lihasten supistumista, atropiinia käytetään lääkkeenä, joka lievittää niiden kouristuksia. (spasmodinen). Näin ollen on tunnettua atropiinin käyttö silmälihasten rentouttamiseen silmänpohjaa tarkasteltaessa sekä kouristuksen lievittämiseen maha-suolikanavan koliikkissa. M-kolinergisiä reseptoreita on myös keskushermostossa (CNS), joten suuret atropiiniannokset voivat aiheuttaa keskushermoston ei-toivotun reaktion: motorista ja henkistä kiihtyneisyyttä, hallusinaatioita, kouristuksia.
Atropiinissa on rakenteeltaan asetyylikoliinin kaltaisia funktionaalisia ryhmiä ja niiden tilajärjestelyjä, joten se kuuluu M-kolinergisten reseptorien kilpaileviin estäjiin. Koska asetyylikoliinin sitoutuminen M-kolinergisiin reseptoreihin aiheuttaa sileiden lihasten supistumista, atropiinia käytetään lääkkeenä, joka lievittää niiden kouristuksia. (spasmodinen). Näin ollen on tunnettua atropiinin käyttö silmälihasten rentouttamiseen silmänpohjaa tarkasteltaessa sekä kouristuksen lievittämiseen maha-suolikanavan koliikkissa. M-kolinergisiä reseptoreita on myös keskushermostossa (CNS), joten suuret atropiiniannokset voivat aiheuttaa keskushermoston ei-toivotun reaktion: motorista ja henkistä kiihtyneisyyttä, hallusinaatioita, kouristuksia.
Ditiliini on H-kolinergisten reseptorien kilpaileva agonisti, joka estää hermo-lihassynapsien toimintaa.
 Luustolihasten neuromuskulaariset synapsit sisältävät H-kolinergisiä reseptoreita. Niiden vuorovaikutus asetyylikoliinin kanssa johtaa lihasten supistuksiin. Joissakin kirurgisissa leikkauksissa sekä endoskooppisissa tutkimuksissa käytetään lääkkeitä, jotka aiheuttavat luurankolihasten rentoutumista. (lihasrelaksantit). Näitä ovat dityliini, joka on asetyylikoliinin rakenteellinen analogi. Se kiinnittyy H-kolinergisiin reseptoreihin, mutta toisin kuin asetyylikoliini, tuhoaa sen hyvin hitaasti. Ionikanavien pitkittyneen avautumisen ja kalvon jatkuvan depolarisaation seurauksena hermoimpulssin johtuminen häiriintyy ja lihasten rentoutuminen tapahtuu. Alun perin nämä ominaisuudet löydettiin curare-myrkystä, joten tällaisia lääkkeitä kutsutaan curariform.
Luustolihasten neuromuskulaariset synapsit sisältävät H-kolinergisiä reseptoreita. Niiden vuorovaikutus asetyylikoliinin kanssa johtaa lihasten supistuksiin. Joissakin kirurgisissa leikkauksissa sekä endoskooppisissa tutkimuksissa käytetään lääkkeitä, jotka aiheuttavat luurankolihasten rentoutumista. (lihasrelaksantit). Näitä ovat dityliini, joka on asetyylikoliinin rakenteellinen analogi. Se kiinnittyy H-kolinergisiin reseptoreihin, mutta toisin kuin asetyylikoliini, tuhoaa sen hyvin hitaasti. Ionikanavien pitkittyneen avautumisen ja kalvon jatkuvan depolarisaation seurauksena hermoimpulssin johtuminen häiriintyy ja lihasten rentoutuminen tapahtuu. Alun perin nämä ominaisuudet löydettiin curare-myrkystä, joten tällaisia lääkkeitä kutsutaan curariform.
AIHE 1.3. PROTEIINIEN DENATUROINTI JA NIIDEN SPONTAANIN RENATIVOIMISEN MAHDOLLISUUS
1. Koska proteiinien natiivi konformaatio säilyy heikkojen vuorovaikutusten vuoksi, proteiinia ympäröivän ympäristön koostumuksen ja ominaisuuksien muutokset, kemiallisten reagenssien ja fysikaalisten tekijöiden vaikutus aiheuttaa muutoksen niiden konformaatiossa (konformaatiolabiteetin ominaisuus). Suuren määrän sidosten katkeaminen johtaa alkuperäisen konformaation tuhoutumiseen ja proteiinin denaturoitumiseen.
Proteiinin denaturaatio- tämä on niiden luonnollisen konformaation tuhoamista denaturoivien aineiden vaikutuksesta, mikä johtuu heikkojen sidosten katkeamisesta, jotka stabiloivat proteiinin avaruudellista rakennetta. Denaturoitumiseen liittyy proteiinin ainutlaatuisen kolmiulotteisen rakenteen ja aktiivisen keskuksen tuhoutuminen ja sen biologisen aktiivisuuden menetys (kuva 1.16).
Kaikki yhden proteiinin denaturoidut molekyylit saavat satunnaisen konformaation, joka eroaa saman proteiinin muista molekyyleistä. Aktiivisen keskuksen muodostavat aminohapporadikaalit osoittautuvat avaruudellisesti etäämmiksi toisistaan, ts. proteiinin spesifinen sitoutumiskohta ligandin kanssa tuhoutuu. Denaturaation aikana proteiinien primäärirakenne pysyy muuttumattomana.
Denaturointiaineiden käyttö biologisessa tutkimuksessa ja lääketieteessä. Biokemiallisissa tutkimuksissa proteiinit poistetaan yleensä ensin liuoksesta ennen alhaisen molekyylipainon yhdisteiden määritystä biologisesta materiaalista. Tähän tarkoitukseen käytetään useimmiten trikloorietikkahappoa (TCA). Kun TCA on lisätty liuokseen, denaturoidut proteiinit saostuvat ja ne poistetaan helposti suodattamalla (taulukko 1.1.)
Lääketieteessä denaturoivia aineita käytetään usein lääketieteellisten instrumenttien ja materiaalien sterilointiin autoklaaveissa (denaturoiva aine - korkea lämpötila) ja antiseptisinä aineina (alkoholi, fenoli, kloramiini) patogeenista mikroflooraa sisältävien saastuneiden pintojen käsittelyyn.
2. Spontaani proteiinin uusiutuminen- todiste proteiinien primäärirakenteen, konformaation ja toiminnan determinismistä. Yksittäiset proteiinit ovat yhden geenin tuotteita, joilla on identtinen aminohapposekvenssi ja jotka saavat saman konformaation solussa. Peruspäätelmä, että proteiinin primäärirakenne sisältää jo tietoa sen konformaatiosta ja toiminnasta, tehtiin joidenkin proteiinien (erityisesti ribonukleaasin ja myoglobiinin) kyvyn perusteella spontaaniin renativaatioon - niiden alkuperäisen konformaation palautumiseen denaturoinnin jälkeen.
Proteiinin avaruudellisten rakenteiden muodostuminen tapahtuu itsekokoamismenetelmällä - spontaanilla prosessilla, jossa polypeptidiketjulla, jolla on ainutlaatuinen primäärirakenne, on taipumus omaksua konformaatio, jossa on alhaisin vapaaenergia liuoksessa. Kyky regeneroida proteiineja, jotka säilyttävät primäärirakenteensa denaturoinnin jälkeen, kuvattiin kokeessa ribonukleaasientsyymillä.
Ribonukleaasi on entsyymi, joka katkaisee sidoksia yksittäisten nukleotidien välillä RNA-molekyylissä. Tällä pallomaisella proteiinilla on yksi polypeptidiketju, jonka tertiääristä rakennetta stabiloivat monet heikot ja neljä disulfidisidosta.
Ribonukleaasin käsittely urealla, joka katkaisee vetysidoksia molekyylissä, ja pelkistimellä, joka katkaisee disulfidisidoksia, johtaa entsyymin denaturoitumiseen ja sen aktiivisuuden menettämiseen.
Denaturoivien aineiden poistaminen dialyysillä johtaa proteiinin konformaation ja toiminnan palautumiseen, ts. reanimaatioon. (Kuva 1.17).
 Riisi. 1.17. Ribonukleaasin denaturaatio ja renaturaatio
Riisi. 1.17. Ribonukleaasin denaturaatio ja renaturaatio
A - ribonukleaasin natiivi konformaatio, jonka tertiäärisessä rakenteessa on neljä disulfidisidosta; B - denaturoitu ribonukleaasimolekyyli;
B - Renatiivinen ribonukleaasimolekyyli, jolla on palautettu rakenne ja toiminta
1. Täytä taulukko 1.2.
Taulukko 1.2. Aminohappojen luokitus radikaalien polariteetin mukaan
2. Kirjoita tetrapeptidin kaava:
Asp - Pro - Fen - Liz
a) eristetään peptidin toistuvat ryhmät, jotka muodostavat peptidirungon, ja vaihtelevat ryhmät, joita edustavat aminohapporadikaalit;
b) osoittavat N- ja C-päät;
c) alleviivaa peptidisidokset;
d) kirjoittaa toinen peptidi, joka koostuu samoista aminohapoista;
e) laskea mahdollisten tetrapeptidivarianttien lukumäärä, joilla on samanlainen aminohappokoostumus.
3. Selitä proteiinien primäärirakenteen rooli nisäkkään neurohypofyysin kahden rakenteellisesti samanlaisen ja evoluutionaalisesti läheisen peptidihormonin - oksitosiinin ja vasopressiinin - vertailevan analyysin esimerkillä (taulukko 1.3).
Taulukko 1.3. Oksitosiinin ja vasopressiinin rakenne ja toiminta
 Tätä varten:
Tätä varten:
a) vertaa näiden kahden peptidin koostumusta ja aminohapposekvenssiä;
b) löytää näiden kahden peptidin primäärirakenteen samankaltaisuus ja niiden biologisen vaikutuksen samankaltaisuus;
c) löytää erot näiden kahden peptidin rakenteessa ja erot niiden toiminnassa;
d) tehdä johtopäätös peptidien primäärirakenteen vaikutuksesta niiden toimintoihin.
4. Kuvaa globulaaristen proteiinien konformaation muodostumisen päävaiheet (sekundaariset, tertiaariset rakenteet, supersekundaarisen rakenteen käsite). Määritä proteiinirakenteiden muodostumiseen osallistuvien sidostyypit. Mitkä aminohapporadikaalit voivat osallistua hydrofobisten vuorovaikutusten, ionisten, vetysidosten muodostumiseen.
Antaa esimerkkejä.
5. Määrittele käsite "proteiinien konformationaalinen labilisuus", osoita sen olemassaolon ja merkityksen syyt.
6. Selitä seuraavan lauseen merkitys: "Proteiinit toimivat niiden spesifisen vuorovaikutuksen perusteella ligandin kanssa", käyttämällä termejä ja selittäkää niiden merkitys: proteiinin konformaatio, aktiivinen kohta, ligandi, komplementaarisuus, proteiinin toiminta.
7. Selitä yhden esimerkin avulla, mitä domeenit ovat ja mikä niiden rooli proteiinien toiminnassa on.
ITSEHALLINTA TEHTÄVÄT
1. Aseta ottelu.
Funktionaalinen ryhmä aminohapporadikaalissa:
A. Karboksyyliryhmä B. Hydroksyyliryhmä C Guanidiiniryhmä D. Tioliryhmä E. Aminoryhmä
2. Valitse oikeat vastaukset.
Aminohapot, joissa on polaarisia varautumattomia radikaaleja, ovat:
A. Tsis B. Asn
B. Glu G. Kolme
3. Valitse oikeat vastaukset.
Aminohapporadikaalit:
A. Tarjoa primaarirakenteen spesifisyys B. Osallistu tertiaarisen rakenteen muodostukseen
B. Koska ne sijaitsevat proteiinin pinnalla, ne vaikuttavat sen liukoisuuteen D. Muodostavat aktiivisen keskuksen
D. Osallistu peptidisidosten muodostukseen
4. Valitse oikeat vastaukset.
Hydrofobisia vuorovaikutuksia voi muodostua aminohapporadikaalien välillä:
A. Tre Lay B. Pro Three
B. Met Ile G. Tir Ala D. Val Fen
5. Valitse oikeat vastaukset.
Ionisidoksia voi muodostua aminohapporadikaalien välille:
A. Gln Asp B. Apr Liz
B. Liz Glu G. Hanhet Asp D. Asn Apr
6. Valitse oikeat vastaukset.
Aminohapporadikaalien välille voi muodostua vetysidoksia:
A. Ser Gln B. Cis Tre
B. Asp Liz G. Glu Asp D. Asn Tre
7. Aseta ottelu.
Proteiinirakenteen muodostumiseen osallistuvan sidoksen tyyppi:
A. Ensisijainen rakenne B. Toissijainen rakenne
B. Tertiäärinen rakenne
D. Ylisekundaarinen rakenne E. Konformaatio.
1. Peptidirungon atomien väliset vetysidokset
2. Heikot sidokset aminohapporadikaalien funktionaalisten ryhmien välillä
3. Sidot aminohappojen α-amino- ja α-karboksyyliryhmien välillä
8. Valitse oikeat vastaukset. Trypsiini:
A. Proteolyyttinen entsyymi B. Sisältää kaksi domeenia
B. Hydrolysoi tärkkelystä
D. Aktiivinen keskus sijaitsee verkkotunnusten välissä. D. Koostuu kahdesta polypeptidiketjusta.
9. Valitse oikeat vastaukset. Atropiini:
A. Neurotransmitteri
B. Asetyylikoliinin rakenteellinen analogi
B. Vuorovaikuttaa H-kolinergisten reseptorien kanssa
G. Tehostaa hermoimpulssin johtumista kolinergisten synapsien läpi
D. M-kolinergisten reseptorien kilpaileva estäjä
10. Valitse oikeat väitteet. Proteiinissa:
V. Ensisijainen rakenne sisältää tietoa aktiivisen sivustonsa rakenteesta
B. Aktiivinen keskus muodostuu primäärirakenteen tasolle
B. Konformaatio on jäykästi kiinnitetty kovalenttisilla sidoksilla
D. Aktiivinen kohta voi olla vuorovaikutuksessa samanlaisten ligandien ryhmän kanssa
johtuu proteiinien D konformationaalisesta labilisuudesta. Ympäristön muuttaminen voi vaikuttaa aktiivisen proteiinin affiniteettiin
keskustasta ligandiin
1. 1-C, 2-D, 3-B.
3. A, B, C, D.
7. 1-B, 2-D, 3-A.
8. A, B, C, D.
PERUSKÄSITTEET JA KÄSITTEET
1. Proteiini, polypeptidi, aminohapot
2. Primaariset, sekundaariset, tertiääriset proteiinirakenteet
3. Konformaatio, natiivi proteiinin konformaatio
4. Kovalenttiset ja heikot sidokset proteiinissa
5. Konformaatiolabiteetti
6. Proteiinin aktiivinen kohta
7. Ligandit
8. Proteiinin taittaminen
9. Ligandien rakenteelliset analogit
10. Domain-proteiinit
11. Yksinkertaiset ja monimutkaiset proteiinit
12. Proteiinien denaturointi, denaturointiaineet
13. Proteiinin uudistaminen
Ratkaista ongelmia
"Proteiinien rakenneorganisaatio ja niiden toiminnan perusta"
1. Proteiinin - hemoglobiini A (HbA) - päätehtävä on hapen kuljettaminen kudoksiin. Ihmispopulaatiossa tunnetaan tämän proteiinin useita muotoja, joilla on muuttuneet ominaisuudet ja toiminta - niin sanotut epänormaalit hemoglobiinit. Esimerkiksi sirppisoluanemiaa (HbS) sairastavien potilaiden punasoluista löydetyn hemoglobiini S:n on havaittu olevan alhainen liukoisuus olosuhteissa, joissa hapen osapaine on alhainen (kuten tapahtuu laskimoveressä). Tämä johtaa tämän proteiinin aggregaattien muodostumiseen. Proteiini menettää toimintansa, saostuu ja punasolut muuttuvat epäsäännöllisiksi (jotkut niistä muodostavat sirpin muodon) ja tuhoutuvat tavallista nopeammin pernassa. Seurauksena kehittyy sirppisoluanemia.
Ainoa ero HvA:n primäärirakenteessa havaittiin hemoglobiinin β-ketjun N-pään alueella. Vertaa β-ketjun N-terminaalisia alueita ja näytä kuinka muutokset proteiinin primäärirakenteessa vaikuttavat sen ominaisuuksiin ja toimintoihin.
 Tätä varten:
Tätä varten:
a) Kirjoita aminohappokaavat, joilla HvA eroaa, ja vertaa näiden aminohappojen ominaisuuksia (polariteetti, varaus).
b) tehdä johtopäätös liukoisuuden vähenemisen ja kudoksen hapen kuljetuksen häiriintymisen syystä.
2. Kuvassa on kaavio proteiinin rakenteesta, jossa on ligandia sitova keskus (aktiivinen keskus). Selitä, miksi proteiini on valikoiva ligandin valinnassa. Tätä varten:
a) muista mikä on proteiinin aktiivinen keskus ja harkitse kuvassa näkyvän proteiinin aktiivisen keskuksen rakennetta;
b) kirjoittaa aktiivisen keskuksen muodostavien aminohapporadikaalien kaavat;
c) piirtää ligandi, joka voisi spesifisesti olla vuorovaikutuksessa proteiinin aktiivisen kohdan kanssa. Merkitse siihen funktionaaliset ryhmät, jotka pystyvät muodostamaan sidoksia aminohapporadikaalien kanssa, jotka muodostavat aktiivisen keskuksen;
d) osoittavat sidostyypit, joita syntyy ligandin ja aktiivisen keskuksen aminohapporadikaalien välillä;
e) Selitä proteiinin ja ligandin vuorovaikutuksen spesifisyyden perusteet.
 3.
Kuvassa näkyy proteiinin aktiivinen kohta ja useita ligandeja.
3.
Kuvassa näkyy proteiinin aktiivinen kohta ja useita ligandeja.
Määritä, mikä ligandeista on todennäköisimmin vuorovaikutuksessa proteiinin aktiivisen kohdan kanssa ja miksi.
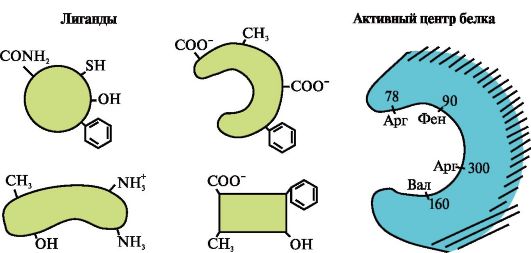 Millaisia sidoksia syntyy proteiini-ligandikompleksin muodostumisen aikana?
Millaisia sidoksia syntyy proteiini-ligandikompleksin muodostumisen aikana?
4. Luonnollisten proteiiniligandien rakenteellisia analogeja voidaan käyttää lääkkeinä proteiinien aktiivisuuden muuttamiseksi.
Asetyylikoliini on hermo-lihassynapsien viritysvälityksen välittäjä. Kun asetyylikoliini on vuorovaikutuksessa proteiinien kanssa - luurankolihasten postsynaptisen kalvon reseptorit, ionikanavat avautuvat ja lihasten supistuminen tapahtuu. Dityliini on lääke, jota käytetään joissakin leikkauksissa lihasten rentouttamiseen, koska se häiritsee hermoimpulssien siirtymistä neuromuskulaaristen synapsien kautta. Selitä dityliinin vaikutusmekanismi lihasrelaksanttina. Tätä varten:
a) kirjoittaa asetyylikoliinin ja dityliinin kaavat ja vertailla niiden rakenteita;
b) kuvaile dityliinin rentouttavan vaikutuksen mekanismia.
5. Joissakin sairauksissa potilaan ruumiinlämpö nousee, mitä pidetään kehon suojaavana reaktiona. Korkeat lämpötilat ovat kuitenkin haitallisia kehon proteiineille. Selitä, miksi yli 40 °C:n lämpötiloissa proteiinien toiminta häiriintyy ja ihmishenkiä uhkaa. Muista tehdäksesi tämän:
1) Proteiinien rakenne ja sidokset, jotka pitävät sen rakenteen alkuperäisessä konformaatiossa;
2) Miten proteiinien rakenne ja toiminta muuttuvat lämpötilan noustessa?;
3) Mitä on homeostaasi ja miksi ihmisten terveyden ylläpitäminen on tärkeää.
Moduuliyksikkö 2 OLIGOMEERINEN PROTEIINIT SÄÄNTELYN VAIKUTUKSEN KOHTEENA. RAKENNE JA TOIMINNALLINEN MONIMENETEÄ PROTEIINEJA. PROTEIINIEN EROTUS- JA PUHDISTUSMENETELMÄT
Oppimistavoitteet Kykyä:
1. Käytä tietoa oligomeeristen proteiinien rakenteen ja toiminnan ominaisuuksista ymmärtääksesi niiden toimintojen säätelyn adaptiivisia mekanismeja.
2. Selitä kaperonien rooli solun proteiinikonformaation synteesissä ja ylläpitämisessä.
3. Selitä elämän ilmenemismuotojen monimuotoisuus elimistössä syntetisoitujen proteiinien rakenteiden ja toimintojen monimuotoisuudella.
4. Analysoi proteiinien rakenteen ja toiminnan välistä suhdetta vertaamalla toisiinsa liittyviä hemoproteiineja - myoglobiinia ja hemoglobiinia sekä immunoglobuliiniperheen viiden proteiiniluokan edustajia.
5. Hyödynnä tietoa proteiinien fysikaalis-kemiallisten ominaisuuksien piirteistä valitaksesi menetelmät niiden puhdistamiseksi muista proteiineista ja epäpuhtauksista.
6. Tulkitse veriplasman proteiinien kvantitatiivisen ja laadullisen koostumuksen tuloksia kliinisen diagnoosin vahvistamiseksi tai selkeyttämiseksi.
Tietää:
1. Oligomeeristen proteiinien rakenteen piirteet ja niiden toimintojen säätelyn adaptiiviset mekanismit hemoglobiinin esimerkissä.
2. Chaperonien rakenne ja toiminnot sekä niiden merkitys proteiinien natiivin konformaation ylläpitämisessä solussa.
3. Periaatteet proteiinien ryhmittelystä perheisiin niiden konformaation ja toimintojen samankaltaisuuden mukaan immunoglobuliinien esimerkissä.
4. Menetelmät proteiinien erottamiseksi niiden fysikaalis-kemiallisten ominaisuuksien perusteella.
5. Veriplasman elektroforeesi menetelmänä proteiinien laadullisen ja kvantitatiivisen koostumuksen arvioimiseksi.
AIHE 1.4. OLIGOMEERISTEN PROTEIINIEN RAKENTEEN JA TOIMINNAN OMINAISUUDET ESIMERKKI HEMOGLOBIINISSA
1. Monet proteiinit sisältävät useita polypeptidiketjuja. Tällaisia proteiineja kutsutaan oligomeerinen, ja yksittäisiä piirejä protomeerit. Oligomeeristen proteiinien protomeerit on yhdistetty monilla heikoilla ei-kovalenttisilla sidoksilla (hydrofobinen, ioninen, vety). Vuorovaikutus
protomeerit suoritetaan ansiosta täydentävyyttä niiden kosketuspinnat.
Oligomeeristen proteiinien protomeerien määrä voi vaihdella suuresti: hemoglobiini sisältää 4 protomeeriä, aspa- 12 protomeeriä ja tupakan mosaiikkiviruksen proteiini sisältää 2120 protomeeriä, jotka on yhdistetty ei-kovalenttisilla sidoksilla. Siksi oligomeeristen proteiinien molekyylipainot voivat olla erittäin korkeat.
Yhden protomeerin vuorovaikutusta muiden kanssa voidaan pitää erikoistapauksena proteiinin vuorovaikutuksesta ligandin kanssa, koska jokainen protomeeri toimii ligandina muille protomeereille. Proteiinissa olevien protomeerien lukumäärää ja yhdistämismenetelmää kutsutaan kvaternäärinen proteiinirakenne.
Proteiinit voivat sisältää protomeerejä, joilla on sama tai erilainen rakenne, esimerkiksi homodimeerit ovat proteiineja, jotka sisältävät kaksi identtistä protomeeriä, ja heterodimeerit ovat proteiineja, jotka sisältävät kaksi eri protomeeriä.
Jos proteiinit sisältävät erilaisia protomeerejä, niihin voi muodostua sitoutumiskeskuksia erilaisilla ligandeilla, jotka ovat rakenteeltaan erilaisia. Kun ligandi sitoutuu aktiiviseen keskukseen, tämän proteiinin toiminta ilmenee. Eri protomeerissä sijaitsevaa keskustaa kutsutaan allosteeriseksi (muuksi kuin aktiiviseksi). Yhteydenotto allosteerinen ligandi tai efektori, se suorittaa säätelytoiminnon (kuva 1.18). Allosteerisen keskuksen vuorovaikutus efektorin kanssa aiheuttaa konformaatiomuutoksia koko oligomeerisen proteiinin rakenteessa sen konformationaalisuuden vuoksi. Tämä vaikuttaa aktiivisen kohdan affiniteettiin spesifiseen ligandiin ja säätelee kyseisen proteiinin toimintaa. Muutosta kaikkien protomeerien konformaatiossa ja toiminnassa oligomeerisen proteiinin vuorovaikutuksessa vähintään yhden ligandin kanssa kutsutaan yhteistoiminnalliseksi konformaatiomuutokseksi. Proteiinin toimintaa tehostavia efektejä kutsutaan aktivaattorit ja efektorit, jotka heikentävät sen toimintaa - estäjät.
Siten oligomeerisissä proteiineissa sekä proteiineissa, joilla on domeenirakenne, ilmenee uusi ominaisuus verrattuna monomeerisiin proteiineihin - kyky allosteeriseen toimintojen säätelyyn (säätely liittämällä proteiiniin erilaisia ligandeja). Tämä voidaan nähdä vertaamalla kahden läheisesti sukua olevan monimutkaisen proteiinin, myoglobiinin ja hemoglobiinin, rakenteita ja toimintoja.
 Riisi. 1.18. Kaavio dimeerisen proteiinin rakenteesta
Riisi. 1.18. Kaavio dimeerisen proteiinin rakenteesta
2. Avaruusrakenteiden muodostuminen ja myoglobiinin toiminta.
Myoglobiini (Mb) on punaisissa lihaksissa esiintyvä proteiini, jonka päätehtävänä on luoda intensiiviseen lihastyöhön tarvittavia O 2 -varastoja. MB on monimutkainen proteiini, joka sisältää proteiiniosan - apoMB:n ja ei-proteiiniosan - hemin. ApoMB:n primäärirakenne määrää sen kompaktin pallomaisen konformaation ja aktiivisen keskuksen rakenteen, johon myoglobiinin ei-proteiiniosa, hemi, on kiinnittynyt. Verestä lihaksiin tuleva happi sitoutuu myoglobiinin koostumuksessa olevaan Fe + 2 -hemiin. MB on monomeerinen proteiini, jolla on erittäin korkea affiniteetti O 2:een, joten myoglobiini vapauttaa happea vain intensiivisen lihastyön aikana, kun O 2:n osapaine laskee jyrkästi.
Konformaation MB muodostuminen. Punaisissa lihaksissa, ribosomeissa translaation aikana, tapahtuu MB:n primäärirakenteen synteesi, jota edustaa spesifinen 153 aminohappotähteen sekvenssi. Mv:n toissijainen rakenne sisältää kahdeksan α-heliksiä, joita kutsutaan latinalaisiksi kirjaimilla A:sta H:hen, joiden välissä on spiraalittomia osia. Mv:n tertiäärinen rakenne on muodoltaan kompakti pallo, jonka syvennyksessä F- ja E α-heliksien välissä on aktiivinen keskus (kuva 1.19).
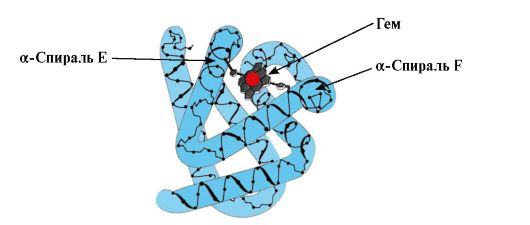 Riisi. 1.19. Myoglobiinin rakenne
Riisi. 1.19. Myoglobiinin rakenne
3. MV-aktiivisen keskuksen rakenteen ja toiminnan ominaisuudet. Mv:n aktiivinen keskus muodostuu pääasiassa hydrofobisista aminohapporadikaaleista, jotka ovat kaukana toisistaan primäärirakenteessa (esim. 3 9 ja Phen 138) Veteen huonosti liukenevat ligandit, hemi ja 02, ovat kiinnittyneet aktiiviseen keskustaan. Hemi on spesifinen apoMv-ligandi (kuva 1.20), joka perustuu neljään pyrrolirenkaaseen, jotka on yhdistetty metenyylisilloilla; keskellä on Fe+ 2 -atomi, joka on yhdistetty pyrrolirenkaiden typpiatomeihin neljällä koordinaatiosidoksella. Aminohappojen hydrofobisten radikaalien lisäksi Mv:n aktiivinen keskus sisältää myös kahden aminohapon tähteitä, joissa on hydrofiilisiä radikaaleja - Gis E 7(Gis 64) ja Gis F8(Hänen 93) (Kuva 1.21).
 Riisi. 1.20. Hemin rakenne - myoglobiinin ja hemoglobiinin ei-proteiiniosa
Riisi. 1.20. Hemin rakenne - myoglobiinin ja hemoglobiinin ei-proteiiniosa
 Riisi. 1.21. Hemin ja O 2:n sijainti apomyoglobiinin ja hemoglobiinin protomeerien aktiivisessa kohdassa
Riisi. 1.21. Hemin ja O 2:n sijainti apomyoglobiinin ja hemoglobiinin protomeerien aktiivisessa kohdassa
Hemi on kovalenttisesti sitoutunut His F 8:aan rautaatomin kautta. O 2 kiinnittyy rautaan hemitason toisella puolella. Hänen E 7 on välttämätön O 2:n oikealle suuntaukselle ja helpottaa hapen lisäämistä Fe + 2 -heemiin
Gis F8 muodostaa koordinaatiosidoksen Fe+ 2:n kanssa ja kiinnittää hemin tiukasti aktiiviseen kohtaan. Gis E 7 on tarpeen oikean suuntautumisen kannalta toisen ligandin - O 2:n aktiivisessa keskustassa sen vuorovaikutuksessa Fe + 2 -hemin kanssa. Hemimikroympäristö luo olosuhteet O 2:n vahvalle mutta palautuvalle sitoutumiselle Fe + 2:n kanssa ja estää vettä pääsemästä hydrofobiseen aktiiviseen keskukseen, mikä voi johtaa sen hapettumiseen Fe + 3 :ksi.
MB:n monomeerinen rakenne ja sen aktiivinen keskus määrää proteiinin korkean affiniteetin 02:lle.
4. Hb:n oligomeerinen rakenne ja Hb-affiniteetin säätely 02:lle ligandeilla. Ihmisen hemoglobiinit- proteiiniperhe sekä monimutkaisiin proteiineihin (hemoproteiineihin) liittyvä myoglobiini. Niillä on tetrameerinen rakenne ja ne sisältävät kaksi a-ketjua, mutta eroavat kahden muun polypeptidiketjun (2a-, 2x-ketjut) rakenteesta. Toisen polypeptidiketjun rakenne määrittää näiden Hb-muotojen toiminnan piirteet. Noin 98 % aikuisen punasolujen hemoglobiinista on hemoglobiini A(2a-, 2p-ketjut).
Sikiön kehityksen aikana hemoglobiineja on kahta päätyyppiä: alkion HB(2α, 2ε), joka löytyy sikiön kehityksen alkuvaiheista, ja hemoglobiini F (sikiö)- (2α, 2γ), joka korvaa varhaisen sikiön hemoglobiinin kuudennen sikiön kehityskuukauden aikana ja korvataan Hb A:lla vasta syntymän jälkeen.
Hv A on myoglobiiniin (Mv) liittyvä proteiini, jota löytyy aikuisten punasoluista. Sen yksittäisten protomeerien rakenne on samanlainen kuin myoglobiinin. Myoglobiinin ja hemoglobiinin protomeerien toissijaiset ja tertiaariset rakenteet ovat hyvin samankaltaisia huolimatta siitä, että vain 24 aminohappotähdettä ovat identtisiä niiden polypeptidiketjujen primäärirakenteessa (hemoglobiinin protomeerien sekundaarirakenne, kuten myoglobiini, sisältää kahdeksan α-heliksiä, merkitty latinalaisilla kirjaimilla A:sta H:hen, ja tertiäärinen rakenne on tiiviin globulin muotoinen). Mutta toisin kuin myoglobiinilla, hemoglobiinilla on oligomeerinen rakenne, se koostuu neljästä polypeptidiketjusta, jotka on yhdistetty ei-kovalenttisilla sidoksilla (kuva 1.22).
Jokainen Hb-protomeeri liittyy ei-proteiiniosaan - hemiin ja viereisiin protomeereihin. Hb:n proteiiniosan yhteys hemiin on samanlainen kuin myoglobiinin: proteiinin aktiivisessa keskustassa hemin hydrofobisia osia ympäröivät hydrofobiset aminohapporadikaalit, paitsi His F 8 ja His E 7 , jotka sijaitsevat molemmilla puolilla hemitasoa ja joilla on samanlainen rooli proteiinin toiminnassa ja sen sitoutumisessa happeen (katso myoglobiinin rakenne).
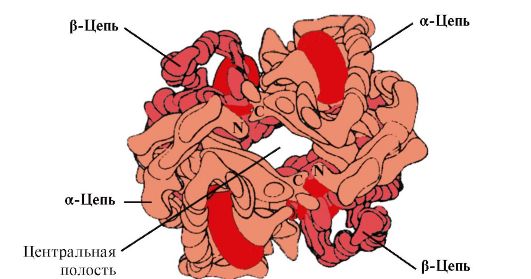 Riisi. 1.22. Hemoglobiinin oligomeerinen rakenne
Riisi. 1.22. Hemoglobiinin oligomeerinen rakenne
Sitä paitsi, Gis E 7 suorittaa tärkeän lisärooli NV:n toiminnassa. Vapaalla heemillä on 25 000 kertaa suurempi affiniteetti CO:lle kuin O 2:lle. CO:ta muodostuu elimistössä pieniä määriä, ja sen korkean affiniteetin hemiin vuoksi se voi häiritä solujen elämälle välttämättömän O 2:n kuljetusta. Hemoglobiinin koostumuksessa hemin affiniteetti hiilimonoksidiin ylittää kuitenkin affiniteetin O 2:lle vain 200 kertaa, koska His:n aktiivisessa keskustassa on E7. Tämän aminohapon jäännös luo optimaaliset olosuhteet hemin sitoutumiselle O2:een ja heikentää hemin vuorovaikutusta CO:n kanssa.
5. Hb:n päätehtävä on O 2:n kuljettaminen keuhkoista kudoksiin. Toisin kuin monomeerinen myoglobiini, jolla on erittäin korkea affiniteetti O 2:een ja joka varastoi happea punaisiin lihaksiin, hemoglobiinin oligomeerinen rakenne tarjoaa:
1) Hb:n nopea kyllästyminen hapella keuhkoissa;
2) Hb:n kyky vapauttaa happea kudoksissa suhteellisen korkeassa 02:n osapaineessa (20-40 mm Hg);
3) mahdollisuus säädellä Hb:n affiniteettia 02:een.
6. Yhteistoiminnalliset muutokset hemoglobiinin protomeerien konformaatiossa nopeuttavat O 2:n sitoutumista keuhkoissa ja sen palautumista kudoksiin. Keuhkoissa korkea 02:n osapaine edistää sen sitoutumista Hb:hen neljän protomeerin (2α ja 2β) aktiivisessa kohdassa. Kunkin protomeerin aktiivinen keskus, kuten myoglobiinissa, sijaitsee kahden α-kierteen (F ja E) välissä hydrofobisessa taskussa. Se sisältää ei-proteiiniosan - hemin, joka on kiinnittynyt proteiiniosaan monilla heikkoilla hydrofobisilla vuorovaikutuksilla ja yhden vahvan sidoksen Fe 2 + -hemin ja His F 8:n välillä (katso kuva 1.21).
Deoksihemoglobiinissa, johtuen tästä yhteydestä His F 8:aan, Fe 2+ -atomi työntyy esiin hemitasosta kohti histidiiniä. O 2:n sitoutuminen Fe 2+:aan tapahtuu hemin toisella puolella His E 7 -alueella yhden vapaan koordinaatiosidoksen avulla. His E 7 tarjoaa optimaaliset olosuhteet O 2:n sitoutumiselle hemiraudan kanssa.
O 2:n lisäys yhden protomeerin Fe +2 -atomiin saa sen siirtymään hemitasoon ja sen taakse siihen liittyvän histidiinitähteen
 Riisi. 1.23. Muutos hemoglobiinin protomeerin konformaatiossa yhdistettynä O 2:een
Riisi. 1.23. Muutos hemoglobiinin protomeerin konformaatiossa yhdistettynä O 2:een
Tämä johtaa muutokseen kaikkien polypeptidiketjujen konformaatiossa niiden konformationaalisuuden vuoksi. Muiden ketjujen konformaation muuttaminen helpottaa niiden vuorovaikutusta seuraavien O 2 -molekyylien kanssa.
Neljäs O 2 -molekyyli kiinnittyy hemoglobiiniin 300 kertaa helpommin kuin ensimmäinen (kuva 1.24).
 Riisi. 1.24. Yhteistoiminnalliset muutokset hemoglobiinin protomeerien konformaatiossa sen vuorovaikutuksessa O 2:n kanssa
Riisi. 1.24. Yhteistoiminnalliset muutokset hemoglobiinin protomeerien konformaatiossa sen vuorovaikutuksessa O 2:n kanssa
Kudoksissa jokainen myöhempi O 2 -molekyyli lohkeutuu helpommin kuin edellinen, myös protomeerikonformaation yhteistoiminnallisten muutosten vuoksi.
7. CO 2 ja H + muodostuvat katabolian aikana eloperäinen aine, vähentävät hemoglobiinin affiniteettia O 2:een suhteessa niiden pitoisuuteen. Solujen toimintaan tarvittava energia tuotetaan pääasiassa mitokondrioissa orgaanisten aineiden hapettumisen aikana hemoglobiinin keuhkoista kuljettaman O 2:n avulla. Orgaanisten aineiden hapettumisen seurauksena muodostuu niiden hajoamisen lopputuotteita: CO 2 ja K 2 O, joiden määrä on verrannollinen käynnissä olevien hapetusprosessien intensiteettiin.
CO 2 diffundoituu soluista vereen ja tunkeutuu punasoluihin, joissa se muuttuu karbanhydraasientsyymin vaikutuksesta hiilihapoksi. Tämä heikko happo hajoaa protoniksi ja bikarbonaatti-ioniksi.
H+ voivat liittyä GIS-radikaaleihin 14 6 hemoglobiinin a- ja p-ketjuissa, so. alueilla, jotka ovat kaukana hemistä. Hemoglobiinin protonoituminen vähentää sen affiniteettia O 2:een, edistää O 2:n poistumista oksiHb:stä, deoksiHb:n muodostumista ja lisää kudosten hapen saantia suhteessa muodostuneiden protonien määrään (kuva 1.25).
Vapautuneen hapen määrän lisääntymistä punasolujen H + -pitoisuuden kasvusta riippuen kutsutaan Bohrin efektiksi (tanskalaisen fysiologin Christian Bohrin mukaan, joka löysi tämän vaikutuksen ensimmäisenä).
Hapen korkea osapaine keuhkoissa edistää sen sitoutumista deoksiHb:hen, mikä vähentää proteiinin affiniteettia H+:aan. Karbanhydraasin vaikutuksesta vapautuneet protonit vuorovaikuttavat bikarbonaattien kanssa muodostaen CO 2:ta ja H 2 O:ta
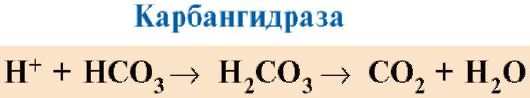
 Riisi. 1.25. Hb:n 02-affiniteetin riippuvuus CO 2:n ja protonien pitoisuudesta (Bohr-ilmiö):
Riisi. 1.25. Hb:n 02-affiniteetin riippuvuus CO 2:n ja protonien pitoisuudesta (Bohr-ilmiö):
A- CO2- ja H+-pitoisuuden vaikutus 02:n vapautumiseen Hb-kompleksista (Bohr-ilmiö); B- deoksihemoglobiinin hapetus keuhkoissa, hiilidioksidin muodostuminen ja vapautuminen.
Tuloksena oleva CO 2 tulee keuhkorakkuloihin ja poistuu uloshengitetyn ilman mukana. Siten hemoglobiinin vapauttaman hapen määrää kudoksissa säätelevät orgaanisten aineiden hajoamistuotteet: mitä voimakkaammin aineet hajoavat esimerkiksi fyysisen rasituksen aikana, sitä korkeampi on CO 2 - ja H + -pitoisuus ja sitä enemmän kudokset saavat happea, koska H:n affiniteetti O 2:een vähenee.
8. Hb-affiniteetin allosteerinen säätely 02:lle ligandilla - 2,3-bisfosfoglyseraatilla. Punasoluissa hemoglobiinin allosteerinen ligandi, 2,3-bisfosfoglyseraatti (2,3-BPG), syntetisoidaan glukoosin hapettumistuotteesta - 1,3-bisfosfoglyseraatista. SISÄÄN normaaleissa olosuhteissa 2,3-BPG:n pitoisuus on korkea ja verrattavissa Hb-pitoisuuteen. 2,3-BPG:llä on vahva negatiivinen varaus -5.
 Kudoskapillaareissa oleva bisfosfoglyseraatti sitoutumalla deoksihemoglobiiniin lisää hapen tuotantoa kudoksissa vähentäen Hb:n affiniteettia O 2:een.
Kudoskapillaareissa oleva bisfosfoglyseraatti sitoutumalla deoksihemoglobiiniin lisää hapen tuotantoa kudoksissa vähentäen Hb:n affiniteettia O 2:een.
Tetrameerisen hemoglobiinimolekyylin keskellä on ontelo. Se muodostuu kaikkien neljän protomeerin aminohappotähteistä (katso kuva 1.22). Kudoskapillaareissa Hb:n protonoituminen (Bohr-ilmiö) katkaisee sidoksen hemiraudan ja O 2:n välillä. Molekyylissä
deoksihemoglobiini, verrattuna oksihemoglobiiniin, ilmaantuu ylimääräisiä ionisidoksia, jotka yhdistävät protomeerit, minkä seurauksena keskusontelon koko kasvaa verrattuna oksihemoglobiiniin. Keskusontelo on paikka, jossa 2,3-BPG kiinnittyy hemoglobiiniin. Keskusontelon koon eroista johtuen 2,3-BPG voi kiinnittyä vain deoksihemoglobiiniin.
2,3-BPG on vuorovaikutuksessa hemoglobiinin kanssa alueella, joka on kaukana proteiinin aktiivisista kohdista ja kuuluu allosteerinen(säätely) ligandit, ja keskusontelo Hb on allosteerinen keskus. 2,3-BPG:llä on voimakas negatiivinen varaus ja se on vuorovaikutuksessa kahden Hb-β-ketjun viiden positiivisesti varautuneen ryhmän kanssa: N-terminaalinen α-aminoryhmä Val ja Lys 82 Gis 143 -radikaalit (kuva 1.26).
 Riisi. 1.26. BPG deoksihemoglobiinin keskusontelossa
Riisi. 1.26. BPG deoksihemoglobiinin keskusontelossa
BPG sitoutuu kolmeen positiivisesti varautuneeseen ryhmään kussakin β-juosteessa.
Kudoskapillaareissa tuloksena oleva deoksihemoglobiini on vuorovaikutuksessa 2,3-BPG:n kanssa, ja p-ketjujen positiivisesti varautuneiden radikaalien ja negatiivisesti varautuneen ligandin välille muodostuu ionisidoksia, jotka muuttavat proteiinin konformaatiota ja vähentävät Hb:n affiniteettia O 2:lle. Hb:n affiniteetin heikkeneminen 02:lle edistää O 2:n tehokkaampaa vapautumista kudokseen.
Keuhkoissa korkeassa osapaineessa happi vuorovaikuttaa Hb:n kanssa liittyen hemiraudaan; tässä tapauksessa proteiinin konformaatio muuttuu, keskusontelo pienenee ja 2,3-BPG siirtyy allosteerisesta keskustasta
Siten oligomeerisillä proteiineilla on uusia ominaisuuksia verrattuna monomeerisiin proteiineihin. ligandien kiinnittäminen paikkoihin,
avaruudellisesti etäällä toisistaan (allosteerinen), jotka voivat aiheuttaa konformaatiomuutoksia koko proteiinimolekyylissä. Vuorovaikutuksesta säätelevien ligandien kanssa proteiinimolekyylin konformaatio muuttuu ja toiminta mukautuu ympäristön muutoksiin.
AIHE 1.5. PROTEIINIEN ALKUMUUN SÄILYTTÄMINEN SOLU-OLLOISSA
Soluissa polypeptidiketjujen synteesin aikana niiden kuljettaminen kalvojen läpi solun vastaaviin osiin, laskostumisprosessissa (natiivin konformaation muodostuminen) ja oligomeeristen proteiinien kokoamisen aikana sekä niiden toiminnan aikana, välituote , aggregaatioalttiita, epästabiileja konformaatioita syntyy proteiinirakenteessa. Hydrofobiset radikaalit, jotka ovat tavallisesti piilossa proteiinimolekyylin sisällä luonnollisessa konformaatiossaan, ilmestyvät pinnalle epävakaassa konformaatiossa ja pyrkivät yhdistymään muiden proteiinien ryhmiin, jotka ovat samoin huonosti veteen liukenevia. Kaikkien tunnettujen organismien soluista on löydetty erityisiä proteiineja, jotka mahdollistavat soluproteiinien optimaalisen laskostumisen, stabiloivat niiden natiivia konformaatiota toiminnan aikana ja mikä tärkeintä, ylläpitävät solunsisäisten proteiinien rakennetta ja toimintoja homeostaasihäiriön sattuessa. Näitä proteiineja kutsutaan "seuraajat" joka tarkoittaa ranskaksi lastenhoitajaa.
1. Molekyylikaperonit ja niiden rooli proteiinien denaturoitumisen estämisessä.
Chaperonit (III) luokitellaan alayksiköiden massan mukaan. Suuren molekyylipainon omaavien chaperonien massa on 60-110 kD. Niistä kolmea luokkaa on tutkittu eniten: Sh-60, Sh-70 ja Sh-90. Jokainen luokka sisältää sukulaisia proteiineja. Siten Sh-70 sisältää proteiineja, joiden molekyylipaino on 66 - 78 kD. Pienen molekyylipainon omaavien chaperonien molekyylipaino on 40-15 kD.
Ohjaajien joukossa on konstitutiivinen proteiineja, joiden korkea perussynteesi ei riipu kehon soluihin kohdistuvista stressaavista vaikutuksista, ja indusoituva, jonka synteesi normaaleissa olosuhteissa on heikkoa, mutta lisääntyy jyrkästi stressaavissa vaikutuksissa. Indusoituvia chaperoneja kutsutaan myös "lämpösokkiproteiineiksi", koska ne löydettiin ensimmäisen kerran korkeille lämpötiloille altistetuista soluista. Soluissa proteiinien suuren pitoisuuden vuoksi osittain denaturoituneiden proteiinien spontaani regeneraatio on vaikeaa. Sh-70 voi estää alkanutta denaturaatioprosessia ja auttaa palauttamaan proteiinien alkuperäisen konformaation. Molekyyliset chaperonit-70- erittäin konservoitunut proteiiniluokka, jota löytyy solun kaikista osista: sytoplasmasta, tumasta, endoplasmisesta retikulumista, mitokondrioista. Sh-70:n ainoan polypeptidiketjun karboksyylipäässä on alue, joka on ura, joka voi olla vuorovaikutuksessa pitkien peptidien kanssa.
7 - 9 aminohappotähdettä, jotka on rikastettu hydrofobisilla radikaaleilla. Tällaisia kohtia pallomaisissa proteiineissa esiintyy suunnilleen joka 16. aminohappo. Sh-70 pystyvät suojaamaan proteiineja lämpöinaktivaatiolta ja palauttamaan osittain denaturoituneiden proteiinien konformaation ja aktiivisuuden.
2. Chaperonien rooli proteiinin laskostuksessa. Proteiinien synteesin aikana ribosomissa polypeptidin N-terminaalinen alue syntetisoidaan ennen C-terminaalista aluetta. Proteiinin täydellinen aminohapposekvenssi tarvitaan luonnollisen konformaation muodostamiseen. Proteiinisynteesiprosessissa chaperones-70 pystyvät aktiivisen keskuksensa rakenteesta johtuen peittämään aggregaatiolle alttiita polypeptidikohtia, jotka ovat rikastuneet hydrofobisilla aminohapporadikaaleilla, kunnes synteesi on valmis (Kuva 1.27, A).
 Riisi. 1.27. Chaperonien osallistuminen proteiinien laskostumiseen
Riisi. 1.27. Chaperonien osallistuminen proteiinien laskostumiseen
A - chaperons-70:n osallistuminen hydrofobisten vuorovaikutusten ehkäisyyn syntetisoidun polypeptidin paikkojen välillä; B - natiiviproteiinin konformaation muodostuminen chaperonikompleksissa
Monet suuren molekyylipainon proteiinit, joilla on monimutkainen konformaatio, kuten domeenirakenne, laskostuvat W-60:n muodostamaan erityiseen tilaan. Sh-60 toimivat oligomeerisena kompleksina, joka koostuu 14 alayksiköstä. Ne muodostavat kaksi onttoa rengasta, joista jokainen koostuu seitsemästä alayksiköstä, nämä renkaat ovat yhteydessä toisiinsa. Jokainen III-60:n alayksikkö koostuu kolmesta domeenista: apikaalinen (apikaalinen), rikastettu renkaan onteloa päin olevilla hydrofobisilla radikaaleilla, väli- ja ekvatoriaalinen (kuva 1.28).
 Riisi. 1.28. 14 Sh-60:stä koostuvan chaperoniinikompleksin rakenne
Riisi. 1.28. 14 Sh-60:stä koostuvan chaperoniinikompleksin rakenne
A - sivukuva; B - ylhäältä katsottuna
Syntetisoidut proteiinit, joissa on laskostumattomille molekyyleille ominaisia pintaelementtejä, erityisesti hydrofobisia radikaaleja, tulevat chaperonirenkaiden onteloon. SISÄÄN erityinen ympäristö näistä onteloista etsitään mahdollisia konformaatioita, kunnes löydetään ainoa, energeettisesti edullisin (kuva 1.27, B). Konformaatioiden muodostumiseen ja proteiinin vapautumiseen liittyy ATP-hydrolyysi ekvatoriaalisella alueella. Tyypillisesti tällainen chaperone-riippuvainen taittaminen vaatii huomattavan määrän energiaa.
Sen lisäksi, että chaperone osallistuu proteiinien kolmiulotteisen rakenteen muodostukseen ja osittain denaturoituneiden proteiinien renativoimiseen, niitä tarvitaan myös sellaisiin perusprosesseihin kuin oligomeeristen proteiinien kokoamiseen, denaturoituneiden proteiinien tunnistamiseen ja kuljettamiseen lysosomeihin, proteiinien kuljettamiseen. kalvojen kautta ja osallistuminen proteiinikompleksien toiminnan säätelyyn.
AIHE 1.6. ERILAISIA PROTEIINEJA. PROTEIINIPERHEET ESIMERKKIIN IMMUNOGLOBULIINEISTA
1. Proteiineilla on ratkaiseva rooli yksittäisten solujen ja koko monisoluisen organismin elämässä, ja niiden toiminnot ovat yllättävän monipuoliset. Tämän määräävät proteiinien primäärirakenteen ja konformaatioiden erityispiirteet, aktiivisen keskuksen ainutlaatuinen rakenne ja kyky sitoa spesifisiä ligandeja.
Vain hyvin pieni osa kaikista mahdollisista peptidiketjujen muunnelmista voi omaksua vakaan tilarakenteen; suurin osa
niistä voi olla monia konformaatioita suunnilleen samalla Gibbsin energialla, mutta eri ominaisuuksilla. Useimpien tunnettujen proteiinien primäärirakenne, joka on valittu biologisen evoluution perusteella, tarjoaa yhdelle konformaatiosta poikkeuksellisen stabiilisuuden, mikä määrää tämän proteiinin toiminnan ominaisuudet.
2. Proteiiniperheet. Yhden sisällä lajit aminohappotähteiden substituutiot voivat johtaa erilaisten proteiinien syntymiseen, jotka suorittavat toisiinsa liittyviä tehtäviä ja joilla on homologiset aminohapposekvenssit. Tällaisilla sukulaisproteiineilla on hämmästyttävän samankaltaiset konformaatiot: a-heliksien ja/tai p-rakenteiden lukumäärä ja järjestys ja useimmat polypeptidiketjujen käännökset ja laskokset ovat samanlaisia tai identtisiä. Proteiinit, joilla on homologisia polypeptidiketjun alueita, samanlainen konformaatio ja vastaavat toiminnot, eristetään proteiiniperheiksi. Esimerkkejä proteiiniperheistä: seriiniproteinaasit, immunoglobuliiniperhe, myoglobiiniperhe.
Seriiniproteinaasit- proteiiniperhe, joka suorittaa proteolyyttisten entsyymien tehtävää. Näitä ovat ruoansulatusentsyymit - kymotrypsiini, trypsiini, elastaasi ja monet veren hyytymistekijät. Näissä proteiineissa on 40 % identtisiä aminohappoja ja hyvin samanlainen konformaatio (kuva 1.29).

Riisi. 1.29. Elastaasin (A) ja kymotrypsiinin (B) spatiaaliset rakenteet
Jotkut aminohapposubstituutiot ovat johtaneet muutokseen näiden proteiinien substraattispesifisyydessä ja toiminnallisen monimuotoisuuden syntymiseen perheen sisällä.
3. Immunoglobuliinien perhe. Kolmeen proteiiniperheeseen kuuluvan immunoglobuliinien superperheen proteiineilla on valtava rooli immuunijärjestelmän toiminnassa:
Vasta-aineet (immunoglobuliinit);
T-lymfosyyttireseptorit;
Tärkeimmän proteiinit - MHC 1. ja 2. luokka (Major Histocompatibility Complex).
Kaikilla näillä proteiineilla on domeenirakenne, ne koostuvat homologisista immuunityyppisistä domeeneista ja suorittavat samanlaisia tehtäviä: ne ovat vuorovaikutuksessa vieraiden rakenteiden kanssa, joko liuenneena vereen, imusolmukkeisiin tai solujen väliseen nesteeseen (vasta-aineet) tai solujen pinnalla (omissa tai ulkomaalainen).
4. Vasta-aineet- B-lymfosyyttien tuottamat spesifiset proteiinit vasteena vieraan rakenteen nielemiseen antigeeni.
Vasta-aineiden rakenteen ominaisuudet
Yksinkertaisimmat vasta-ainemolekyylit koostuvat neljästä polypeptidiketjusta: kahdesta identtisestä kevyestä ketjusta - L, joka sisältää noin 220 aminohappoa, ja kahdesta identtisestä raskaasta ketjusta - H, joka koostuu 440-700 aminohaposta. Kaikki neljä vasta-ainemolekyylin ketjua on yhdistetty useilla ei-kovalenttisilla sidoksilla ja neljällä disulfidisidoksella (kuva 1.30).
Vasta-aineiden kevytketjut koostuvat kahdesta domeenista: variaabeli (VL), joka sijaitsee polypeptidiketjun N-terminaalisella alueella, ja vakio (CL), joka sijaitsee C-päässä. Raskaissa ketjuissa on tyypillisesti neljä domeenia: yksi muuttuja (VH) N-päässä ja kolme vakiota (CH1, CH2, CH3) (katso kuva 1.30). Jokaisella immunoglobuliinidomeenilla on β-laskostettu ylärakenne, jossa kaksi kysteiinitähdettä on liitetty toisiinsa disulfidisidoksella.
Kahden vakiodomeenin CH1 ja CH2 välissä on alue, joka sisältää suuren määrän proliinitähteitä, jotka estävät sekundaarirakenteen muodostumisen ja viereisten H-ketjujen vuorovaikutuksen tässä segmentissä. Tämä sarana-alue antaa vasta-ainemolekyylille joustavuuden. Raskaan ja kevyen ketjun vaihtelevien domeenien välissä on kaksi identtistä antigeeniä sitovaa kohtaa (aktiivisia kohtia antigeenien sitomiselle), joten tällaisia vasta-aineita kutsutaan usein ns. bivalentteja. Antigeenin sitoutuminen vasta-aineeseen ei koske molempien ketjujen vaihtelevien alueiden koko aminohapposekvenssiä, vaan vain 20-30 aminohappoa, jotka sijaitsevat kunkin ketjun hypervariaabelialueilla. Juuri nämä alueet määrittävät kunkin vasta-ainetyypin ainutlaatuisen kyvyn olla vuorovaikutuksessa vastaavan komplementaarisen antigeenin kanssa.
Vasta-aineet ovat yksi kehon puolustuslinjoista tunkeutuvia vieraita organismeja vastaan. Niiden toiminta voidaan jakaa kahteen vaiheeseen: ensimmäinen vaihe on antigeenin tunnistaminen ja sitoutuminen vieraiden organismien pinnalle, mikä on mahdollista, koska vasta-ainerakenteessa on antigeeniä sitovia kohtia; toinen vaihe on antigeenin inaktivaatio- ja tuhoutumisprosessin käynnistäminen. Toisen vaiheen spesifisyys riippuu vasta-aineluokasta. Raskasketjuja on viisi luokkaa, jotka eroavat toisistaan vakiodomeenien rakenteessa: α, δ, ε, γ ja μ, joiden mukaan erotetaan viisi immunoglobuliiniluokkaa: A, D, E, G ja M.
Raskaiden ketjujen rakenteelliset ominaisuudet antavat raskaiden ketjujen sarana-alueille ja C-terminaalisille alueille kullekin luokalle ominaisen konformaation. Kun antigeeni sitoutuu vasta-aineeseen, konformaatiomuutokset vakiodomeeneissa määrittävät polun antigeenin poistamiseen.
 Riisi. 1. 30. IgG:n domeenirakenne
Riisi. 1. 30. IgG:n domeenirakenne
Immunoglobuliinit M
Immunoglobuliineilla M on kaksi muotoa.
Monomeerinen muoto- kehittyvän B-lymfosyytin tuottamien vasta-aineiden 1. luokka. Myöhemmin monet B-solut siirtyvät tuottamaan muita vasta-aineluokkia, mutta joilla on sama antigeenia sitova kohta. IgM liitetään kalvoon ja toimii antigeenin tunnistavana reseptorina. IgM:n sisällyttäminen solukalvoon on mahdollista, koska alueen häntäosassa on 25 hydrofobista aminohappotähdettä.
IgM:n eritysmuoto sisältää viisi monomeerista alayksikköä, jotka on kytketty toisiinsa disulfidisidoksilla, ja lisäksi polypeptidin J-ketjun (kuva 1.31). Tämän muodon raskasketjuiset monomeerit eivät sisällä hydrofobista häntää. Pentameerissä on 10 antigeeniä sitovaa kohtaa, ja siksi se tunnistaa ja poistaa tehokkaasti antigeenin, joka on päässyt kehoon ensimmäistä kertaa. IgM:n eritysmuoto on pääasiallinen vasta-aineiden luokka, joka erittyy vereen primaarisen immuunivasteen aikana. IgM:n sitoutuminen antigeeniin muuttaa IgM:n konformaatiota ja indusoi sen sitoutumisen komplementtijärjestelmän ensimmäiseen proteiinikomponenttiin (komplementtijärjestelmä on joukko proteiineja, jotka osallistuvat antigeenin tuhoamiseen) ja tämän järjestelmän aktivoitumisen. Jos antigeeni sijaitsee mikro-organismin pinnalla, komplementtijärjestelmä aiheuttaa solukalvon eheyden rikkoutumisen ja bakteerisolun kuoleman.
Immunoglobuliinit G
Määrällisesti tämä immunoglobuliiniluokka on vallitseva veressä (75 % kaikesta Ig:stä). IgG - monomeerit, pääluokka vasta-aineita, jotka erittyvät vereen sekundaarisen immuunivasteen aikana. IgG:n vuorovaikutuksen jälkeen mikro-organismien pinta-antigeenien kanssa antigeeni-vasta-ainekompleksi pystyy sitomaan ja aktivoimaan komplementtijärjestelmän proteiineja tai voi olla vuorovaikutuksessa makrofagien ja neutrofiilien spesifisten reseptorien kanssa. vuorovaikutus fagosyyttien kanssa
 Riisi. 1.31. IgM:n erittävän muodon rakenne
Riisi. 1.31. IgM:n erittävän muodon rakenne
antigeeni-vasta-ainekompleksien imeytymiseen ja niiden tuhoutumiseen solujen fagosomeissa. IgG on ainoa vasta-aineluokka, joka voi läpäistä istukan esteen ja suojata sikiötä infektioilta kohdussa.
Immunoglobuliinit A
Pääasiallinen vasta-aineiden luokka eritteissä (maito, sylki, hengitysteiden eritteet ja suolistossa). IgA erittyy pääosin dimeerisessä muodossa, jossa monomeerit ovat liittyneet toisiinsa ylimääräisen J-ketjun kautta (kuva 1.32).
IgA ei ole vuorovaikutuksessa komplementtijärjestelmän ja fagosyyttisolujen kanssa, mutta sitoutumalla mikro-organismeihin vasta-aineet estävät niitä kiinnittymästä epiteelisoluihin ja tunkeutumasta kehoon.
Immunoglobuliinit E
Immunoglobuliineja E edustavat monomeerit, joissa raskaat e-ketjut sisältävät, samoin kuin immunoglobuliinien M μ-ketjut, yhden vaihtelevan ja neljä vakiodomeenia. Erityksen jälkeen IgE sitoutuu omaan
 Riisi. 1.32. IgA:n rakenne
Riisi. 1.32. IgA:n rakenne
C-terminaaliset alueet, joissa on vastaavat reseptorit syöttösolujen ja basofiilien pinnalla. Tämän seurauksena niistä tulee näiden solujen pinnalla olevien antigeenien reseptoreita (kuva 1.33).
 Riisi. 1.33. IgE:n vuorovaikutus antigeenin kanssa syöttösolun pinnalla
Riisi. 1.33. IgE:n vuorovaikutus antigeenin kanssa syöttösolun pinnalla
Kun antigeeni on kiinnittynyt vastaaviin antigeeniä sitoviin IgE-kohtiin, solut saavat signaalin biologisesti aktiivisten aineiden (histamiinin, serotoniinin) erittymisestä, jotka ovat suurelta osin vastuussa tulehdusreaktion kehittymisestä ja niiden ilmenemisestä. allergiset reaktiot kuten astma, nokkosihottuma, heinänuha.
Immunoglobuliinit D
Immunoglobuliineja D löytyy seerumista hyvin pieniä määriä, ne ovat monomeerejä. Raskaissa δ-ketjuissa on yksi muuttuva ja kolme vakiodomeenia. IgD toimivat B-lymfosyyttien reseptoreina, muita toimintoja ei vielä tunneta. Spesifisten antigeenien vuorovaikutus B-lymfosyyttien (IgD) pinnalla olevien reseptoreiden kanssa johtaa näiden signaalien välittämiseen soluun ja sellaisten mekanismien aktivoitumiseen, jotka varmistavat tämän lymfosyyttikloonin lisääntymisen.
AIHE 1.7. PROTEIINIEN FYSIKAALISET-KEMIALLISET OMINAISUUDET JA NIIDEN EROTUSMENETELMÄT
1. Yksittäiset proteiinit eroavat fysikaalis-kemiallisista ominaisuuksistaan:
Molekyylien muoto;
Molekyylipaino;
Kokonaisvaraus, jonka arvo riippuu aminohappojen anionisten ja kationisten ryhmien suhteesta;
Polaaristen ja ei-polaaristen aminohapporadikaalien suhde molekyylien pinnalla;
Erilaisten denaturointiaineiden kestävyysasteet.
2. Proteiinien liukoisuus riippuu edellä lueteltujen proteiinien ominaisuuksista sekä väliaineen koostumuksesta, johon proteiini liukenee (pH-arvot, suolakoostumus, lämpötila, muiden orgaanisten aineiden läsnäolo, jotka voivat olla vuorovaikutuksessa proteiinin kanssa). Proteiinimolekyylien varauksen suuruus on yksi niiden liukoisuuteen vaikuttavista tekijöistä. Kun varaus katoaa isoelektrisessä pisteessä, proteiinit aggregoituvat ja saostuvat helpommin. Tämä koskee erityisesti denaturoituja proteiineja, joiden pinnalla on hydrofobisia aminohapporadikaaleja.
Proteiinimolekyylin pinnalla on sekä positiivisesti että negatiivisesti varautuneita aminohapporadikaaleja. Näiden ryhmien lukumäärä ja siten proteiinien kokonaisvaraus riippuu väliaineen pH:sta, ts. H + - ja OH - ryhmien pitoisuuksien suhde. Happamassa ympäristössä H+:n pitoisuuden kasvu johtaa karboksyyliryhmien -COO - + H+ > -COOH dissosioitumisen estymiseen ja proteiinien negatiivisen varauksen vähenemiseen. Alkalisessa ympäristössä aminoryhmien -NH 3 + + OH - - NH 2 + H 2 O hajoamisen aikana muodostuneiden ylimääräisten OH - protonien sitoutuminen veden muodostumisen kanssa johtaa proteiinien positiivisen varauksen vähenemiseen. pH-arvoa, jossa proteiinin nettovaraus on nolla, kutsutaan isoelektrinen piste (IEP). IET:ssä positiivisesti ja negatiivisesti varautuneiden ryhmien määrä on sama, ts. proteiini on isoelektrisessä tilassa.
3. Yksittäisten proteiinien erottaminen. Kehon rakenteen ja toiminnan ominaisuudet riippuvat siinä syntetisoitujen proteiinien joukosta. Proteiinien rakenteen ja ominaisuuksien tutkiminen on mahdotonta ilman niiden eristämistä solusta ja puhdistamista muista proteiineista ja orgaanisista molekyyleistä. Yksittäisten proteiinien eristämisen ja puhdistamisen vaiheet:
solujen tuhoutuminen tutkitusta kudoksesta ja saada homogenaatti.
Homogenaatin erottaminen fraktioiksi sentrifugointi, jolloin saadaan tuma-, mitokondrio-, sytosolinen tai muu fraktio, joka sisältää halutun proteiinin.
Valikoiva lämpödenaturointi- proteiiniliuoksen lyhytaikainen kuumennus, jossa osa denaturoituneista proteiiniepäpuhtauksista voidaan poistaa (jos proteiini on suhteellisen termisesti stabiili).
Suolaus pois. Eri proteiinit saostuvat eri suolapitoisuuksilla liuoksessa. Vähitellen suolapitoisuutta lisäämällä on mahdollista saada useita yksittäisiä fraktioita, joissa yhdessä niistä on vallitseva erittyvän proteiinin pitoisuus. Yleisimmin käytetty proteiinien fraktiointi on ammoniumsulfaatti. Vähiten liukenevat proteiinit saostuvat alhaisilla suolapitoisuuksilla.
Geelisuodatus- menetelmä molekyylien seulomiseksi turvonneiden Sephadex-rakeiden läpi (kolmiulotteiset dekstraanipolysakkaridiketjut, joissa on huokoset). Proteiinien kulkunopeus Sephadexillä täytetyn kolonnin läpi riippuu niiden molekyylipainosta: mitä pienempi proteiinimolekyylien massa on, sitä helpommin ne tunkeutuvat rakeisiin ja pysyvät siellä pidempään, mitä suurempi massa, sitä nopeammin ne eluoituvat rakeista. sarakkeessa.
Ultrasentrifugointi- menetelmä, joka koostuu siitä, että sentrifugiputkessa olevat proteiinit asetetaan ultrasentrifugin roottoriin. Roottorin pyöriessä proteiinien sedimentaationopeus on verrannollinen niiden molekyylipainoon: raskaampien proteiinien fraktiot sijaitsevat lähempänä putken pohjaa, kevyemmät lähempänä pintaa.
elektroforeesi- menetelmä, joka perustuu eroihin proteiinien liikkumisnopeudessa sähkökentässä. Tämä arvo on verrannollinen proteiinien varaukseen. Proteiinielektroforeesi suoritetaan paperilla (tässä tapauksessa proteiinien liikenopeus on verrannollinen vain niiden varaukseen) tai polyakryyliamidigeelissä, jolla on tietty huokoskoko (proteiinien liikenopeus on verrannollinen niiden varaukseen ja molekyylipainoon ).
Ioninvaihtokromatografia- fraktiointimenetelmä, joka perustuu ionisoituneiden proteiiniryhmien sitomiseen vastakkaisesti varautuneisiin ioninvaihtohartsiryhmiin (liukenemattomat polymeerimateriaalit). Proteiinin sitoutumisvoima hartsiin on verrannollinen proteiinin varaukseen. Ioninvaihtopolymeeriin adsorboituneet proteiinit voidaan pestä pois kasvavilla NaCl-liuospitoisuuksilla; mitä pienempi proteiinivaraus, sitä pienempi NaCl-pitoisuus vaaditaan hartsin ioniryhmiin liittyvän proteiinin pesemiseksi pois.
Affiniteettikromatografia- spesifisin menetelmä yksittäisten proteiinien eristämiseksi Proteiinin ligandi kiinnitetään kovalenttisesti inerttiin polymeeriin. Kun proteiiniliuos johdetaan kolonnin läpi polymeerin kanssa, johtuen proteiinin komplementaarisesta sitoutumisesta ligandiin, vain tälle ligandille spesifinen proteiini adsorboituu pylvääseen.
Dialyysi- menetelmä, jota käytetään pienen molekyylipainon yhdisteiden poistamiseen eristetyn proteiinin liuoksesta. Menetelmä perustuu proteiinien kyvyttömyyteen kulkea puoliläpäisevän kalvon läpi, toisin kuin pienimolekyylipainoiset aineet. Sitä käytetään proteiinien puhdistamiseen pienimolekyylisistä epäpuhtauksista, esimerkiksi suoloista suolauksen jälkeen.
TEHTÄVÄT OPIN ULKOPUOLISTA TYÖTÄ
1. Täytä taulukko. 1.4.
Taulukko 1.4. Vertaileva analyysi sukulaisproteiinien - myoglobiinin ja hemoglobiinin - rakenteet ja toiminnot
a) muista aktiivisen keskuksen Mb ja Hb rakenne. Mikä rooli aminohappojen hydrofobisilla radikaaleilla on näiden proteiinien aktiivisten keskusten muodostumisessa? Kuvaa Mb- ja Hb-aktiivisen keskuksen rakenne ja ligandin kiinnittymismekanismit siihen. Mikä rooli His F 8- ja His E 7 -tähteillä on Mv- ja Hv-aktiivisen kohdan toiminnassa?
b) mitä uusia ominaisuuksia monomeeriseen myoglobiiniin verrattuna on läheistä sukua olevalla oligomeeriproteiinilla, hemoglobiinilla? Selitä kooperatiivisten muutosten rooli hemoglobiinimolekyylin protomeerien konformaatiossa, CO 2 - ja protonipitoisuuksien vaikutus hemoglobiinin happiaffiniteettiin ja 2,3-BPG:n rooli Hb-toiminnan allosteerisessa säätelyssä.
2. Kuvaile molekyylikaperonien ominaisuuksia kiinnittäen huomiota niiden rakenteen ja toiminnan väliseen suhteeseen.
3. Mitkä proteiinit on ryhmitelty perheisiin? Määritä tämän perheen proteiinien samanlaiset rakenteelliset ominaisuudet ja niihin liittyvät toiminnot käyttämällä immunoglobuliiniperheen esimerkkiä.
4. Usein biokemiallisiin ja lääketieteellisiin tarkoituksiin tarvitaan puhdistettuja yksittäisiä proteiineja. Selitä, mihin proteiinien fysikaalis-kemiallisiin ominaisuuksiin niiden erottamiseen ja puhdistamiseen käytetyt menetelmät perustuvat.
ITSEHALLINTA TEHTÄVÄT
1. Valitse oikeat vastaukset.
Hemoglobiinin tehtävät:
A. O 2 -kuljetus keuhkoista kudoksiin B. H + -kuljetus kudoksista keuhkoihin
B. Veren pH-arvon ylläpitäminen D. CO2:n kuljetus keuhkoista kudoksiin
D. CO 2:n kuljetus kudoksista keuhkoihin
2. Valitse oikeat vastaukset. ligandiα -Hb-protomeeri on: A. Heme
B. Happi
B. CO D. 2,3-BPG
D. P-protomeeri
3. Valitse oikeat vastaukset.
Hemoglobiini eroaa myoglobiinista:
A. Sillä on kvaternäärinen rakenne
B. Toissijaista rakennetta edustavat vain α-heliksit
B. Viittaa monimutkaisiin proteiineihin
D. Vuorovaikuttaa allosteerisen ligandin kanssa D. Kovalenttisesti sitoutunut hemiin
4. Valitse oikeat vastaukset.
Hb:n affiniteetti O 2:lle laskee:
A. Kun yksi O 2 -molekyyli on kiinnittynyt B. Kun yksi O 2 -molekyyli eliminoituu
B. Kun olet vuorovaikutuksessa 2,3-BPG:n kanssa
D. Kiinnitettynä protomeereihin H + D. Kun 2,3-BPG:n pitoisuus pienenee
5. Aseta ottelu.
Hb-tyypeille on ominaista:
A. Muodostaa fibrillaarisia aggregaatteja deoksimuodossa B. Sisältää kaksi α- ja kaksi δ-ketjua
B. Hb:n vallitseva muoto aikuisten punasoluissa D. Se sisältää hemiä, jonka aktiivisessa keskustassa on Fe + 3
D. Sisältää kaksi α- ja kaksi γ-ketjua 1. HvA 2.
6. Aseta ottelu.
Ligandit Hb:
A. Sitoutuu Hb:hen allosteerisessa keskustassa
B. Sillä on erittäin korkea affiniteetti aktiiviseen kohtaan Hb
B. Liittäminen lisää Hb:n affiniteettia O 2 D:hen. Hapettaa Fe + 2:n Fe + 3:ksi
D. Muodostaa kovalenttisen sidoksen hysF8:n kanssa
7. Valitse oikeat vastaukset.
Saattajat:
A. Proteiinit, joita on solun kaikissa osissa
B. Synteesi tehostuu stressaavissa vaikutuksissa
B. Osallistu denaturoitujen proteiinien hydrolyysiin
D. Osallistu proteiinien luonnollisen konformaation ylläpitämiseen
D. Luo organelleja, joissa muodostuu proteiinikonformaatiota
8. Ottelu. Immunoglobuliinit:
A. Eritysmuoto on pentameerinen
B. Ig-luokka, joka ylittää istukan esteen
B. Ig - syöttösolureseptori
D. Ig:n pääluokka, joka on läsnä epiteelisolujen eritteissä. D. B-lymfosyyttireseptori, jonka aktivointi varmistaa solujen lisääntymisen
9. Valitse oikeat vastaukset.
Immunoglobuliinit E:
A. Makrofagien tuottamat B. Niillä on raskaita ε-ketjuja.
B. Upotettu T-lymfosyyttien kalvoon
D. Toimivat kalvoreseptoreina syöttösolujen ja basofiilien antigeeneille
D. Vastuussa allergisten reaktioiden ilmenemisestä
10. Valitse oikeat vastaukset.
Proteiinien erottelumenetelmä perustuu niiden molekyylipainon eroihin:
A. Geelisuodatus
B. Ultrasentrifugointi
B. PolD. Ioninvaihtokromatografia
D. Affiniteettikromatografia
11. Valitse oikea vastaus.
Proteiinien erottelumenetelmä perustuu eroihin niiden vesiliukoisuudessa:
A. Geelisuodatus B. Suolaus
B. Ioninvaihtokromatografia D. Affiniteettikromatografia
E. Polyakryyliamidigeelielektroforeesi
VASTAUKSIEN STANDARDIT "ITSEENHALLINTA TEHTÄVÄT"
1. A, B, C, D
2. A, B, C, D
5. 1-B, 2-A, 3-D
6. 1-C, 2-B, 3-A
7. A, B, D, D
8. 1-G; 2-B, 3-C
PERUSKÄSITTEET JA KÄSITTEET
1. Oligomeeriset proteiinit, protomeeri, proteiinien kvaternäärinen rakenne
2. Yhteistoiminnalliset muutokset protomeerin konformaatiossa
3. Bohr-efekti
4. Proteiinitoimintojen allosteerinen säätely, allosteerinen keskus ja allosteerinen efektori
5. Molekyylikaperonit, lämpöshokkiproteiinit
6. Proteiiniperheet (seriiniproteaasit, immunoglobuliinit)
7. Rakenteen IgM-, G-, E-, A-yhteys toimintoon
8. Proteiinien kokonaisvaraus, proteiinien isoelektrinen piste
9. Elektroforeesi
10. Suolaus
11. Geelisuodatus
12. Ioninvaihtokromatografia
13. Ultrasentrifugointi
14. Affiniteettikromatografia
15. Plasmaproteiinielektroforeesi
TEHTÄVÄT KUULUTUSTYÖHÄN
1. Vertaa hemoglobiinin (Hb) ja myoglobiinin (Mb) saturaatioasteiden riippuvuutta hapella sen osapaineesta kudoksissa
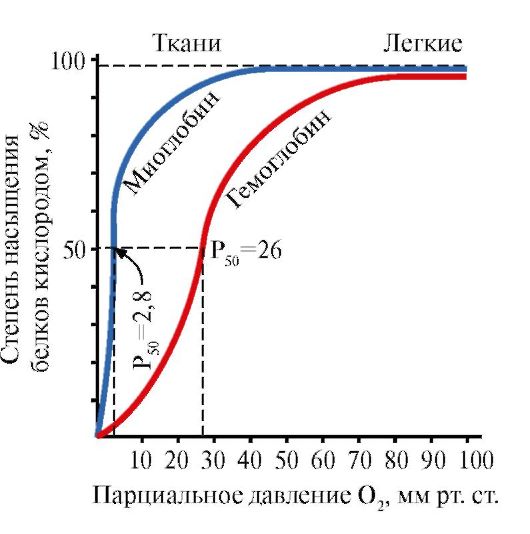 Riisi. 1.34. MV:n jaHbhappea sen osapaineesta
Riisi. 1.34. MV:n jaHbhappea sen osapaineesta
Huomaa, että proteiinin happisaturaatiokäyrien muoto on erilainen: myoglobiinilla - hyperboli, hemoglobiinilla - sigmoidimuoto.
1. Vertaa hapen osapaineen arvoja, joissa Mb ja Hb ovat kyllästyneet O 2:lla 50 %. Millä näistä proteiineista on suurempi affiniteetti O 2:een?
2. Mitkä MB:n rakenteelliset ominaisuudet määräävät sen suuren affiniteetin O 2 :lle?
3. Mitkä Hb:n rakenteelliset ominaisuudet mahdollistavat sen, että se vapauttaa O 2:ta lepäävien kudosten kapillaareista (suhteellisen korkealla O 2:n osapaineella) ja lisää jyrkästi tätä palautusta työskentelevissä lihaksissa? Mikä oligomeeristen proteiinien ominaisuus tarjoaa tämän vaikutuksen?
4. Laske kuinka paljon O 2:ta (%) antaa hapetettua hemoglobiinia lepo- ja työlihakseen?
5. tehdä johtopäätöksiä proteiinin rakenteen ja toiminnan välisestä suhteesta.
2. Hemoglobiinin kapillaareissa vapauttaman hapen määrä riippuu kudosten kataboliaprosessien voimakkuudesta (Bohr-ilmiö). Kuinka muutokset kudosten aineenvaihdunnassa säätelevät Hb:n affiniteettia O 2 :een? CO 2:n ja H+:n vaikutus Hb:n affiniteettiin O 2:een
 1. Kuvaile Bohrin vaikutusta.
1. Kuvaile Bohrin vaikutusta.
2. mihin suuntaan kaaviossa näkyvä prosessi etenee:
a) keuhkojen kapillaareissa;
b) kudoskapillaareissa?
3. Mikä on Bohr-ilmiön fysiologinen merkitys?
4. Miksi Hb:n ja H+:n vuorovaikutus hemistä kaukana olevissa kohdissa muuttaa proteiinin affiniteettia O 2:lle?
3. Hb:n affiniteetti 02:een riippuu sen ligandin, 2,3-bifosfoglyseraatin, pitoisuudesta, joka on Hb:n 02-affiniteetin allosteerinen säätelijä. Miksi ligandin vuorovaikutus aktiivisesta kohdasta kaukana olevassa kohdassa vaikuttaa proteiinin toimintaan? Kuinka 2,3-BPG säätelee Hb:n affiniteettia O 2:een? Voit ratkaista ongelman vastaamalla seuraaviin kysymyksiin:
1. Mistä ja mistä 2,3-bifosfoglyseraattia (2,3-BPG) syntetisoidaan? Kirjoita sen kaava, osoita tämän molekyylin varaus.
2. Minkä muodon hemoglobiinin (oksi tai deoksi) kanssa BPG on vuorovaikutuksessa ja miksi? Millä Hb-molekyylin alueella vuorovaikutus tapahtuu?
3. mihin suuntaan kaaviossa esitetty prosessi etenee?
a) kudosten kapillaareissa;
b) keuhkojen kapillaareissa?
4. missä tulisi olla kompleksin suurin pitoisuus
Nv-2,3-BFG:
a) levossa olevien lihasten kapillaareissa,
b) työskentelevien lihasten kapillaareissa (olettaen, että punasoluissa on sama BPG-pitoisuus)?
5. Miten Hb:n affiniteetti hapen suhteen muuttuu, kun ihminen sopeutuu korkeisiin olosuhteisiin, jos BPG:n pitoisuus punasoluissa kasvaa? Mikä on tämän ilmiön fysiologinen merkitys?
4. 2,3-BPG:n tuhoutuminen säilyneen veren varastoinnin aikana häiritsee Hb:n toimintaa. Miten Hb:n affiniteetti O 2:een muuttuu säilyneessä veressä, jos 2,3-BPG:n pitoisuus punasoluissa voi laskea 8:sta 0,5 mmol/l:aan. Onko mahdollista siirtää tällaista verta vakavasti sairaille potilaille, jos 2,3-BPG-pitoisuus palautuu aikaisintaan kolmen päivän kuluttua? Onko mahdollista palauttaa punasolujen toiminta lisäämällä 2,3-BPG:tä vereen?
5. Muista yksinkertaisimpien immunoglobuliinimolekyylien rakenne. Mikä rooli immunoglobuliineilla on immuunijärjestelmässä? Miksi ig:eitä kutsutaan usein bivalentteiksi? Miten Ig:iden rakenne liittyy niiden toimintaan? (Kuvaile käyttämällä esimerkkiä immunoglobuliiniluokista.)
Proteiinien fysikaalis-kemialliset ominaisuudet ja menetelmät niiden erottamiseksi.
6. Miten proteiinin nettovaraus vaikuttaa sen liukoisuuteen?
a) määritä peptidin kokonaisvaraus pH:ssa 7
Ala-Glu-Tre-Pro-Asp-Liz-Cis
b) kuinka tämän peptidin varaus muuttuu pH:ssa >7, pH<7, рН <<7?
c) mikä on proteiinin isoelektrinen piste (IEP) ja missä ympäristössä se sijaitsee
Tämän peptidin IET?
d) missä pH-arvossa tämän peptidin pienin liukoisuus havaitaan.
7. Miksi piimä, toisin kuin tuore maito, "koaguloituu" keitettäessä (eli kaseiinimaitoproteiini saostuu)? Tuoreen maidon kaseiinimolekyyleillä on negatiivinen varaus.
8. Geelisuodatusta käytetään yksittäisten proteiinien erottamiseen. Seos, joka sisälsi proteiineja A, B ja C, joiden molekyylimassat olivat 160 000, 80 000 ja 60 000, analysoitiin geelisuodatuksella (kuva 1.35). Turvonneet geelirakeet läpäisevät proteiineja, joiden molekyylipaino on alle 70 000. Mikä periaate on tämän erotusmenetelmän taustalla? Mikä kaavioista edustaa oikein fraktioinnin tuloksia? Määritä proteiinien A, B ja C vapautumisjärjestys kolonnista.
 Riisi. 1.35. Geelisuodatusmenetelmän käyttäminen proteiinien erottamiseen
Riisi. 1.35. Geelisuodatusmenetelmän käyttäminen proteiinien erottamiseen
9. Kuvassa 1.36, A esittää kaavion elektroforeesista paperilla terveen ihmisen veren seerumin proteiineista. Tällä menetelmällä saatujen proteiinifraktioiden suhteelliset määrät ovat: albumiinit 54-58 %, a1-globuliinit 6-7 %, a2-globuliinit 8-9 %, p-globuliinit 13 %, y-globuliinit 11-12 %.
 Riisi. 1.36 Terveen ihmisen (A) ja potilaan (B) veriplasmaproteiinien elektroforeesi paperille
Riisi. 1.36 Terveen ihmisen (A) ja potilaan (B) veriplasmaproteiinien elektroforeesi paperille
I - y-globuliinit; II - p-globuliinit; III -α 2 - globuliini; IV-α 2 - globuliini; V - albumiinit
Moniin sairauksiin liittyy kvantitatiivisia muutoksia heraproteiinien koostumuksessa (dysproteinemia). Näiden muutosten luonne otetaan huomioon diagnoosia tehtäessä sekä taudin vaikeusastetta ja vaihetta arvioitaessa.
Käyttämällä taulukossa annettuja tietoja. 1.5, tee oletus sairaudesta, jolle on tunnusomaista kuvassa 1 esitetty elektroforeettinen profiili. 1.36.
Taulukko 1.5. Muutokset veren seerumin proteiinien pitoisuudessa patologiassa



