Sfingomyeliinit. Sfingolipidit, niiden biosynteesi ja biologinen rooli Sfingomyeliinin hydrolyysin aikana niitä muodostuu
2.1.2 Sfingolipidit (sfingofosfolipidit)
Sfingomyeliinit. Nämä ovat yleisimpiä sfingolipidejä. Niitä löytyy pääasiassa eläin- ja kasvisolujen kalvoista. Niitä on erityisen runsaasti hermokudoksessa. Sfingomyeliinejä löytyy myös munuaisten, maksan ja muiden elinten kudoksista. Hydrolyysin aikana sfingomyeliinit muodostavat yhden rasvahappomolekyylin, yhden molekyylin kaksiarvoista tyydyttymätöntä alkoholia, sfingosiinia, yhden molekyylin typpipitoista emästä ja yhden molekyylin fosforihappoa. Sfingomyeliinien yleinen kaava voidaan esittää seuraavasti:
Sfingomyeliinimolekyylin rakentamisen yleinen suunnitelma muistuttaa tietyssä mielessä glyserofosfolipidien rakennetta. Sfingomyeliinimolekyyli sisältää polaarisen "pään", joka sisältää sekä positiivisia (koliinijäännös) että negatiivisia (fosforihappojäännös) varauksia ja kaksi ei-polaarista "häntä" (pitkä alifaattinen sfingosiini- ja rasvahappoasyyliradikaaliketju).
2.2 Glykolipidit (glykosfingolipidit)
Glykolipidejä on laajalti kudoksissa, erityisesti hermokudoksessa, erityisesti aivoissa. Pääasiallinen glykolipidimuoto eläinkudoksissa ovat glykosfingolipidit. Jälkimmäiset sisältävät keramidia, joka koostuu sfingosiinialkoholista ja rasvahappojäännöksestä sekä yhdestä tai useammasta sokeritähteestä.
Yksinkertaisimmat glykosfingolipidit ovat galaktosyylikeramidit ja glukosyylikeramidit.
Galaktosyylikeramidit ovat aivojen ja muiden hermokudosten pääasiallisia sfingolipidejä, mutta niitä löytyy pieninä määrinä myös monissa muissa kudoksissa. Galaktosyylikeramidit sisältävät heksoosia (yleensä D-galaktoosia), joka on liitetty esterisidoksella aminoalkoholin sfingosiinin hydroksyyliryhmään. Lisäksi galaktosyyliseramidi sisältää rasvahappoa. Useimmiten se on lignoseriini-, nervoni- tai serebronihappoa eli 24 hiiliatomia sisältäviä rasvahappoja. On olemassa sulfogalaktosyylikeramideja, jotka eroavat galaktosyylikeramideista siten, että heksoosin kolmanteen hiiliatomiin on kiinnittynyt rikkihappotähde. Nisäkkäiden aivoissa sulfogalaktosyylikeramideja löytyy pääasiassa valkoisesta aineesta, ja niiden tasot aivoissa ovat paljon alhaisemmat kuin galaktosyylikeramidien.
Glukosyyliseramidit ovat yksinkertaisia glykosfingolipidejä, joita esiintyy muissa kudoksissa kuin hermokudoksessa, pääasiassa glukosyylikeramideissa. Niitä on pieniä määriä aivokudoksessa. Toisin kuin galaktosyylikeramideissa, niissä on glukoosijäännös galaktoositähteen sijaan.
Monimutkaisemmat glykosfingolipidit ovat gangliosideja, jotka muodostuvat glykosyylikeramideista. Gangliosidit sisältävät lisäksi yhden tai useamman siaalihappomolekyylin. Ihmiskudoksissa hallitseva siaalihappo on neuramiinihappo. Lisäksi glukoosijäännöksen sijasta ne sisältävät usein kompleksisen oligosakkaridin. Gangliosideja löytyy suuria määriä hermokudoksessa. Ne ilmeisesti suorittavat reseptoreita ja muita toimintoja. Yksi yksinkertaisimmista gangliosideista on gametosidi, joka on eristetty erytrosyyttien stromasta. Se sisältää keramidia, yhden glukoosimolekyylin ja yhden molekyylin N-asetyylineuramiinihappoa.
2.3 Steroidit
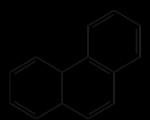
Kaikkia tarkasteltuja lipidejä kutsutaan yleensä saippuoituneiksi, koska niiden alkalinen hydrolyysi tuottaa saippuoita. On kuitenkin olemassa lipidejä, jotka eivät hydrolysoidu vapauttaen rasvahappoja. Nämä lipidit sisältävät steroideja. Steroidit ovat luonnossa laajalle levinneitä yhdisteitä. Ne löytyvät usein yhdessä rasvojen kanssa. Ne voidaan erottaa rasvasta saippuoimalla (ne päätyvät saippuoitumattomaan fraktioon). Kaikilla steroideilla niiden rakenteessa on ydin, joka muodostuu hydratusta fenantreenista (renkaat A, B ja C) ja syklopentaanista (rengas D).



Fenantreeni Perhydrofenantreeni Steroidien yleinen rakenteellinen perusta.
Steroideja ovat esimerkiksi lisämunuaiskuoren hormonit, sappihapot, D-vitamiinit, sydämen glykosidit ja muut yhdisteet. Ihmiskehossa steroidien joukossa tärkeä paikka on steroleilla (steroleilla), eli steroidialkoholeilla. Sterolien pääedustaja on kolesteroli (kolesteroli).
Molekyylin monimutkaisen rakenteen ja epäsymmetrian vuoksi steroideilla on monia mahdollisia stereoisomeerejä. Kukin steroidiytimen kuuden hiilen renkaista (renkaat A, B ja C) voi ottaa kaksi erilaista avaruudellista muotoa - "tuoli" tai "vene" muoto.
Luonnollisissa steroideissa, mukaan lukien kolesteroli, kaikki renkaat ovat "tuolin" muotoisia, mikä on vakaampi rakenne. Renkaat voivat puolestaan olla toisiinsa nähden cis- tai trans-asennossa.
Kolesteroli. Kuten todettiin, steroidien joukossa on ryhmä yhdisteitä, joita kutsutaan steroleiksi (steroleiksi). Steroleille on tunnusomaista hydroksyyliryhmän läsnäolo asemassa 3 sekä sivuketju asemassa 17. Sterolien tärkeimmässä edustajassa, kolesterolissa, kaikki renkaat ovat trans-asemassa; lisäksi siinä on kaksoissidos 5. ja 6. hiiliatomin välillä. Siksi kolesteroli on tyydyttymätön alkoholi. Kolesterolin rengasrakenteelle on ominaista merkittävä jäykkyys, kun taas sivuketju on suhteellisen joustava. Joten kolesteroli sisältää alkoholihydroksyyliryhmän C-3:ssa ja haarautuneen alifaattisen ketjun, jossa on 8 hiiliatomia C-17:ssä. Kolesterolin kemiallinen nimi on 3-hydroksi-5,6-kolestiini. C-3:n hydroksyyliryhmä voidaan esteröidä korkeammalla rasvahapolla, mikä johtaa kolesteroliestereiden (kolesteridien) muodostumiseen.
Kolesteroli on sappihappojen ja steroidihormonien muodostumisen lähde nisäkkäiden kehossa. Kolesterolin fysiologiset toiminnot ovat monipuoliset.





Ympäristönsuojelu Johtopäätös Kuva 2 - Opinnäytetyön verkkokaavio 2.1 Tutkimuskohteet Opinnäytetyön tutkimuskohteena olivat erilaisista luonnonrasvoista eristetyt mikro-organismit: hylje (N), villarasvaalustalla kasvatettu hylje (Hb), villa (W) ) ja mikro-organismeja, jotka on eristetty...
Lipidit sisältävät tavallisten happojen lisäksi ainutlaatuisia mykolihappoja, jotka ovat ominaisia vain näille mikro-organismeille, jotka ovat suurimolekyylisiä b-hydroksihappoja, joilla on pitkä alifaattinen ketju a-asemassa. Rihmasieni-lipidien FCS on suurelta osin identtinen kasviöljyjen koostumuksen kanssa. Tässä suhteessa sienien lipideille voidaan löytää käyttöä kansantalouden eri sektoreilla (...
Vesihaude 10 min. Maitoliemen haju määritetään kuumennettaessa 80-85 asteeseen. Liemen läpinäkyvyysaste määritetään visuaalisesti sylinterissä, jonka halkaisija on 20 mm. Kemiallinen tutkimus. Siipikarjanlihan kemiallisiin tutkimuksiin kuuluvat haihtuvien rasvahappojen määrän määritys, ammoniakin ja ammoniumsuolojen määritys, reaktio peroksidaasille bentsidiinin kanssa, hapon ja peroksidin määritys...





Huomio yllä oleva, on tarkoituksenmukaisempaa ottaa käyttöön teknisiä toimenpiteitä yrityksessä, mikä puolestaan vaikuttaa suotuisasti RUE "Gomel OTKZ -rasvatehtaan" taloudelliseen tulokseen LUKU 3 TAPOJA RAHOITUSTILAN PARANTAMISEKSI RUE:ssa " PUNAINEN BANNERI RASVAKASVIEN GOMELIN TILAUS" 3.1 Toimenpiteen toteuttaminen halvempaa ja vähemmän energistä...
glyserofosfolipidit. Glyserofosfolipidien rakenteellinen perusta on glyseroli. Glyserofosfolipidit ovat molekyylejä, joissa kaksi rasvahappoa on liitetty esterisidoksella glyseroliin ensimmäisessä ja toisessa asemassa; kolmannessa asemassa on fosforihappotähde, johon vuorostaan voidaan kiinnittää erilaisia substituentteja, useimmiten aminoalkoholeja. Jos kolmas asema sisältää vain fosforihappoa, niin glyserofosfolipidiä kutsutaan fosfatidihapoksi. Sen loppuosa on ns "fosfatidyyli"; se sisältyy jäljellä olevien glyserofosfolipidien nimeen, jonka jälkeen on merkitty fosforihapon vetyatomin substituentin nimi, esimerkiksi fosfatidyylietanoliamiini, fosfatidyylikoliini jne. Fosfatidihappoa esiintyy elimistössä pieni määrä vapaana, mutta se on välituote sekä triasyyliglyserolien että glyserofosfolipidien synteesissä. Glyserofosfolipideissä, kuten triasyyliglyseroleissa, toisella sijalla ovat pääasiassa polyeenihapot; kalvorakenteeseen sisältyvässä fosfatidyylikoliinimolekyylissä tämä on useimmiten arakidonihappoa. Kalvofosfolipidien rasvahapot eroavat muista ihmisen lipideistä polyeenihappojen vallitsemisella (jopa 80-85 %), mikä varmistaa kalvorakenteeseen sisältyvien proteiinien toiminnalle välttämättömän hydrofobisen kerroksen nestemäisen tilan.
Glyserofosfolipidien yleinen kaava näyttää tältä:
Toisin kuin fosfatidyylikoliinimolekyylin triglyseridit, yksi glyserolin trihydroksyyliryhmistä ei liity rasvahappoon, vaan fosforihappoon. Lisäksi fosforihappo puolestaan on yhdistetty esterisidoksella typpipitoiseen emäkseen – koliiniin [HO-CH 2 -CH 2 -N + (CH 3) 3]. Siten glyseroli, korkeammat rasvahapot, fosforihappo ja koliini yhdistyvät fosfatidyylikoliinimolekyylissä:

Fosfatidyylietanoliamiinit. Suurin ero fosfatidyylikoliinien ja fosfatidyylietanoliamiinien välillä on typpipitoisen etanoliamiinin (HO-CH 2 -CH 2 -N + H 3) läsnäolo jälkimmäisessä:

Glyserofosfolipideistä fosfatidyylikoliinit ja fosfatidyylietanoliamiinit löytyvät eniten eläinten ja korkeampien kasvien kehosta. Nämä 2 glyserofosfolipidiryhmää ovat metabolisesti sukua keskenään ja ovat solukalvojen tärkeimmät lipidikomponentit.
Fosfatidyyliseriinit. Fosfatidyyliseriinimolekyylissä typpipitoinen yhdiste on seriinin aminohappotähde


Fosfatidyyliseriinit ovat paljon vähemmän yleisiä kuin fosfatidyylikoliinit ja fosfoetanoliamiinit, ja niiden merkityksen määrää pääasiassa se, että ne osallistuvat fosfatidyylietanoliamiinien synteesiin.
Fosfatidyyli-inositolit. Nämä lipidit kuuluvat fosfatidihappojohdannaisten ryhmään, mutta eivät sisällä typpeä. Tämän glyserofosfolipidien alaluokan radikaali (R3) on kuuden hiilen syklinen alkoholiositoli:

Fosfatidyyli-inositolit ovat melko yleisiä luonnossa. Niitä löytyy eläimistä, kasveista ja mikro-organismeista. Eläimillä muutokset havaitaan keuhkojen aivoissa ja maksassa.
Kysymys 36. Sfingolipidit. Rakenne ja rooli.
Sfingolipidit
Aminoalkoholi sfingosiini, joka koostuu 18 hiiliatomista, sisältää hydroksyyliryhmiä ja aminoryhmän. Sfingosiini muodostaa suuren ryhmän lipidejä, joissa rasvahappo on kytketty siihen aminoryhmän kautta. Sfingosiinin ja rasvahapon välisen vuorovaikutuksen tuotetta kutsutaan "keramidi"). Keramideissa rasvahapot kytkeytyvät epätavallisella (amidi)sidoksella ja hydroksyyliryhmät pystyvät olemaan vuorovaikutuksessa muiden radikaalien kanssa. Keramidit erottuvat niiden koostumuksen muodostavista rasvahapporadikaaleista. Tyypillisesti nämä ovat rasvahappoja, joilla on pitkä ketjupituus - 18-26 hiiliatomia. Sfingolipidejä on kolme päätyyppiä:
Keramidit ovat yksinkertaisimpia sfingolipidejä. Ne sisältävät vain sfingosiinia, joka on yhdistetty rasvahappoasyylijäännökseen.
Sfingomyeliinit sisältävät varautuneen polaarisen ryhmän, kuten fosfokoliinin tai fosfoetanoliamiinin.
Glykosfingolipidit sisältävät keramidia, joka on esteröity 1-hydroksiryhmästä sokerijäännöksellä. Sokerista riippuen glykosfingolipidit jaetaan serebrosideihin ja gangliosideihin.
Cerebrosidit sisältävät glukoosia tai galaktoosia sokerijäämänä.
Gangliosidit sisältävät trisakkaridia, joista yksi on aina siaalihappo.
Biol. Sfingolipidien rooli on monipuolinen. Tiedetään, että ne osallistuvat aksonien, synapsien ja muiden hermokudoksen solujen kalvorakenteiden muodostumiseen, välittävät tunnistusmekanismeja, reseptorivuorovaikutuksia, solujen välisiä kontakteja ja muita tärkeitä prosesseja kehossa.
Nämä ovat yleisimpiä sfingolipidejä. Niitä löytyy pääasiassa eläin- ja kasvisolujen kalvoista. Niitä on erityisen runsaasti hermokudoksessa. Sfingomyeliinit löytyvät myös munuaisten, maksan ja muiden elinten kudoksesta. Hydrolyysin aikana sfingomyeliinit muodostavat yhden molekyylin rasvahappoa, yhden molekyylin kaksiarvoista tyydyttymätöntä alkoholia, sfingosiinia, yhden molekyylin typpipitoista emästä (yleensä koliinia) ja yhden molekyylin fosforihappoa. Sfingomyeliinien yleinen kaava voidaan esittää seuraavasti:

Kysymys 37. Glykolipidit esiintyy laajalti kudoksissa, erityisesti hermokudoksessa, erityisesti aivoissa. Pääasiallinen glykolipidimuoto eläinkudoksissa ovat glykosfingolipidit. Jälkimmäiset sisältävät keramidia, joka koostuu sfingosiinialkoholista ja rasvahappojäännöksestä sekä yhdestä tai useammasta sokeritähteestä. Yksinkertaisimmat glykosfingolipidit ovat galaktosyylikeramidit ja glukosyylikeramidit.
Galaktosyylikeramidit ovat aivojen ja muiden hermokudosten pääasiallisia sfingolipidejä, mutta niitä löytyy pieninä määrinä myös monissa muissa kudoksissa. Galaktosyylikeramidit sisältävät heksoosia (yleensä D-galaktoosia), joka on liitetty esterisidoksella aminoalkoholin sfingosiinin hydroksyyliryhmään. Lisäksi galaktosyyliseramidi sisältää rasvahappoa. Useimmiten se on lignoseriini-, nervoni- tai aivohappoa, ts. rasvahapot, joissa on 24 hiiliatomia.

On olemassa sulfogalaktosyylikeramideja, jotka eroavat galaktosyylikeramideista heksoosin kolmanteen hiiliatomiin kiinnittyneen rikkihappojäännöksen läsnäololla. Nisäkkään aivoissa sulfo-galaktosyylikeramideja on pääasiassa valkoisessa aineessa, ja niiden pitoisuus aivoissa on paljon pienempi kuin galaktosyylikeramidien.
Glukosyyliseramidit ovat yksinkertaisia glykosfingolipidejä, joita esiintyy muissa kudoksissa kuin hermokudoksessa ja pääasiassa glukosyylikeramideina. Niitä on pieniä määriä aivokudoksessa. Toisin kuin galaktosyylikeramideissa, niissä on glukoosijäännös galaktoositähteen sijaan. Monimutkaisemmat glykosfingolipidit ovat gangliosideja, jotka muodostuvat glykosyylikeramideista. Gangliosidit sisältävät lisäksi yhden tai useamman siaalihappomolekyylin. Ihmiskudoksissa hallitseva siaalihappo on neuramiinihappo. Lisäksi glukoosijäännöksen sijasta ne sisältävät usein kompleksisen oligosakkaridin. Gangliosideja löytyy suuria määriä hermokudoksessa. Ne ilmeisesti suorittavat reseptoreita ja muita toimintoja. Yksi yksinkertaisimmista gangliosideista on punasolujen stromasta eristetty hematosidi. Se sisältää keramidia (asyylisfingosiinia), yhden molekyylin glukoosia ja yhden molekyylin N-asetyylineuramiinihappoa.

Kysymys38. KOLESTEROLI- tärkeä kalvojen komponentti ja hydrofobisen kerroksen ominaisuuksien säätelijä. Kolesterolijohdannaiset (sappihapot) ovat välttämättömiä rasvojen sulatuksessa. Kolesterolista syntetisoidut steroidihormonit osallistuvat energian, vesi-suola-aineenvaihdunnan ja seksuaalisten toimintojen säätelyyn Ihmiskehossa tämä on päästeroidi, muut steroidit ovat sen johdannaisia. Kasvit, sienet ja hiiva eivät syntetisoi kolesterolia, vaan muodostavat erilaisia fytosteroleja ja mykosteroleja, jotka eivät imeydy ihmiskehoon. Bakteerit eivät pysty syntetisoimaan steroideja. Kolesteroli on osa kalvoja ja vaikuttaa kaksoiskerroksen rakenteeseen ja lisää sen jäykkyyttä. Sappihapot, steroidihormonit ja D3-vitamiini syntetisoidaan kolesterolista. Heikentynyt kolesteroliaineenvaihdunta johtaa ateroskleroosin kehittymiseen. Kolesteroli on molekyyli, joka sisältää 4 fuusioitunutta rengasta, jotka on merkitty latinalaisilla kirjaimilla A, B, C, D, haarautunut sivuketju, jossa on 8 hiiliatomia asemassa 17, 2 "kulmaista" metyyliryhmää (18 ja 19) ja hydroksyyliryhmä asema 3. Rasvahappojen lisääminen esterisidoksella hydroksyyliryhmään johtaa kolesteroliestereiden muodostumiseen. Esteröimättömässä muodossaan kolesteroli on osa eri solujen kalvoja. Kolesterolin hydroksyyliryhmä on vesikerrosta päin, ja molekyylin jäykkä hydrofobinen osa on upotettu kalvon sisäiseen hydrofobiseen kerrokseen. Veressä 2/3 kolesterolista on esteröitynä ja 1/3 vapaana kolesterolina. Kolesteroliesterit toimivat eräissä soluissa (esimerkiksi maksassa, lisämunuaiskuoressa, sukurauhasissa) sen laskeutumismuodossa. Näistä varastoista kolesterolia käytetään sappihappojen ja steroidihormonien synteesiin.
Sfingolipidit. Niitä löytyy pääasiassa eläin- ja kasvisolujen kalvoista. Niitä on erityisen runsaasti hermokudoksessa. Sfingomyeliinit löytyvät myös munuaisten, maksan ja muiden elinten kudoksesta. Hydrolysoituessaan sfingomyeliinit muodostavat yhden molekyylin rasvahappoa, yhden molekyylin kaksiarvoista tyydyttymätöntä alkoholia, sfingosiinia, yhden molekyylin typpipitoista emästä (yleensä koliinia) ja yhden molekyylin fosforihappoa. Sfingomyeliinien yleinen kaava voidaan esittää seuraavasti:
Sfingomyeliinimolekyylin rakentamisen yleinen suunnitelma muistuttaa tietyssä mielessä glyserofosfolipidien rakennetta. Sfingomyeliinimolekyyli sisältää polaarisen "pään", jossa on sekä positiivisia (koliinijäännös) että negatiivisia (fosforihappojäännös) varauksia, ja kaksi ei-polaarista "häntä" (pitkä alifaattinen sfingosiiniketju ja rasvahappoasyyliradikaali). Joistakin sfingomyeliineistä, esimerkiksi niistä, jotka on eristetty aivoista ja pernasta, löydettiin sfingosiinin sijasta alkoholidihydrosfingosiinia (pelkistetty sfingosiini):
7.6 Steroidit
Kaikkia tarkasteltuja lipidejä kutsutaan yleensä saippuoituneiksi, koska niiden alkalinen hydrolyysi tuottaa saippuoita. On kuitenkin olemassa lipidejä, jotka eivät hydrolysoidu vapauttaen rasvahappoja. Nämä lipidit sisältävät steroideja. Steroidit ovat luonnossa laajalle levinneitä yhdisteitä. Ne löytyvät usein yhdessä rasvojen kanssa. Ne voidaan erottaa rasvasta saippuoimalla (ne päätyvät saippuoitumattomaan fraktioon). Kaikilla rakenteessaan olevilla steroideilla on ydin, joka muodostuu hydratusta fenantreenista (renkaat A, B ja C) ja syklopentaanista (rengas D) (kuva 24):

Kuva 24 - Yleistetty steroidiydin
Steroideja ovat esimerkiksi lisämunuaiskuoren hormonit, sappihapot, D-vitamiinit, sydämen glykosidit ja muut yhdisteet. Ihmiskehossa sterolit (sterolit) ovat tärkeässä asemassa steroidien joukossa, ts. steroidiset alkoholit. Sterolien pääedustaja on kolesteroli (kolesteroli).
Molekyylin monimutkaisen rakenteen ja epäsymmetrian vuoksi steroideilla on monia mahdollisia stereoisomeerejä. Kukin steroidiytimen kuuden hiilen renkaista (renkaat A, B ja C) voi ottaa kaksi erilaista avaruudellista muotoa - "tuoli" tai "vene" muoto.
Kolesteroli on sappihappojen sekä steroidihormonien (sukupuoli- ja kortikoidi) muodostumisen lähde nisäkkäiden kehossa. Kolesteroli tai tarkemmin sanottuna sen hapettumistuote - 7-dehydrokolesteroli, muuttuu ihossa UV-säteiden vaikutuksesta D3-vitamiiniksi. Näin ollen kolesterolin fysiologinen toiminta on monipuolinen.
Kolesterolia löytyy eläinrasvoista, mutta ei kasvirasvoista. Kasvit ja hiiva sisältävät rakenteeltaan samanlaisia yhdisteitä kuin kolesteroli, mukaan lukien ergosteroli.

Ergosteroli on D-vitamiinin esiaste. Kun ergosteroli altistuu UV-säteille, sillä on antirakiittisen vaikutuksen ominaisuus (kun B-rengas avautuu).
Kaksoissidoksen palautuminen kolesterolimolekyylissä johtaa koprosterolin (koprostanolin) muodostumiseen. Koprosterolia löytyy ulosteista, ja se muodostuu bakteerien suoliston mikroflooran palauttaessa kolesterolin kaksoissidoksen C5- ja C6-atomien välillä. 
Nämä sterolit, toisin kuin kolesteroli, imeytyvät hyvin huonosti suolistossa, ja siksi niitä löytyy ihmisen kudoksista pieniä määriä.
8 Hiilihydraattien kemia
Termiä "hiilihydraatit" ehdotti ensimmäisenä Dorpatin (nykyisen Tarton) yliopiston professori K.G. Schmidt vuonna 1844. Tuolloin oletettiin, että kaikilla hiilihydraateilla on yleinen kaava C m (H 2 O) n, ts. hiilihydraatti + vesi. Tästä johtuu nimi "hiilihydraatit". Esimerkiksi glukoosilla ja fruktoosilla on kaava C(H2O)6, ruokosokerilla (sakkaroosilla) C12(H2O)11, tärkkelyksellä [C6(H2O)5]n jne. Myöhemmin kävi ilmi, että joukko yhdisteitä, jotka ominaisuuksiltaan kuuluvat hiilihydraattien luokkaan, sisältävät vetyä ja happea hieman eri suhteessa kuin yleisessä kaavassa on ilmoitettu (esimerkiksi deoksiriboosi C 5 H 10 O 4). Vuonna 1927 kansainvälinen kemiannimikkeistön uudistuskomissio ehdotti termin "hiilihydraatit" korvaamista termillä "glysidit", mutta vanha nimi "hiilihydraatit" on juurtunut ja on yleisesti hyväksytty.
Hiilihydraattikemialla on yksi johtavista paikoista orgaanisen kemian kehityksen historiassa. Ruokosokeria voidaan pitää ensimmäisenä kemiallisesti puhtaassa muodossa eristettynä orgaanisena yhdisteenä. Tuotti vuonna 1861 A.M. Butlerovin (kehon ulkopuolella) formaldehydistä muodostuva hiilihydraattien synteesi oli ensimmäinen synteesi, joka edustaa yhtä kolmesta pääaineluokasta (proteiinit, lipidit, hiilihydraatit), jotka muodostavat eläviä organismeja. Yksinkertaisimpien hiilihydraattien kemiallinen rakenne selvitettiin 1800-luvun lopulla. E. Fischerin perustutkimuksen tuloksena. Kotimaiset tutkijat A.A. antoivat merkittävän panoksen hiilihydraattien tutkimukseen. Colley, P.P. Shorygin, N.K. Kochetkov ym. Tämän vuosisadan 20-luvulla englantilaisen tutkijan W. Haworthin työt loivat pohjan polysakkaridien rakennekemialle. 1900-luvun toiselta puoliskolta. Hiilihydraattien kemia ja biokemia kehittyvät nopeasti niiden tärkeän biologisen merkityksen vuoksi.
LIITTOVALTAINEN TERVEYS- JA SOSIAALINEN KEHITYSVIRASTO
Ammattikorkeamman KOULUTUKSEN VALTION OPETUSLAITOS VENÄJÄN VALTION Lääketieteen YLIOPISTO LIITTOVALTION TERVEYS- JA SOSIAALINEN KEHITYSVIRASTO
sfingolipidit.
Niiden biosynteesi ja biologinen rooli
Nikitin Pavel 112 ryhmä
Sfingolipidit ovat ryhmä monimutkaisia lipidejä, joiden molekyylit perustuvat alifaattisiin aminoalkoholeihin, joista yleisimmät ovat sfingosiini ja serebriini.
CH3 (CH2)12CH CH CH CH CH2OH CH3; (CH2)12CH2CHCHCHCH2OH
OH NH2 OH OH NH2
sfingosiini cerebriini (fytosfingosiini)
Sfingolipidit jaetaan kahteen pääryhmään:
Sfingofosfolipidit, jotka sisältävät fosforihapon ja koliinin jäämiä (sfingomyeliinit) tai fosforihappoa ja inosityyliglykosidia (fytosfingolipidit);
sfingoglykolipidit, jotka sisältävät monosakkarideja (yleensä galaktoosia) tai oligosakkarideja (serebrosideja) ; ja siaalihappojäännökset (gangliosidit) .
Sfingomyeliinit ovat runsaimpia sfingolipidejä. Niitä löytyy pääasiassa eläin- ja kasvisolujen kalvoista. Niitä on erityisen runsaasti hermokudoksessa; sfingomyeliinit löytyvät myös munuaisten, maksan ja muiden elinten kudoksista. Sfingomyeliinit muodostavat hydrolyysin aikana yhden rasvahappomolekyylin, yhden rasvahappomolekyylin, yhden kaksiatomisen tyydyttymättömän aminoalkoholin sfingosiinin, yhden molekyylin typpipitoista emästä (yleensä koliinia) ja yhden molekyylin fosforihappoa, minkä vuoksi sph kuuluu siihen. fosfolipidien luokka. Sfingomyeliinien yleinen rakenne näyttää tältä:
Sfingomyeliinimolekyylin konformaatio on joissain suhteissa samanlainen kuin glyserofosfolipidien konformaatio. Sfingomyeliinimolekyyli sisältää polaarisen "pään", jossa on sekä positiivinen (koliinijäännös) että negatiivinen (fosforihappojäännös) varaus, ja kaksi ei-polaarista "häntä" (pitkä alifaattinen sfingosiiniketju ja esteröity rasvahappo). . On huomattava, että joissakin sfingomyeliineissä, esimerkiksi aivoista ja pernasta eristetyissä sfingomyeliineissä, löydettiin alkoholidihydrosfingosiinia (pelkistetty sfingosiini) sfingosiinin sijasta.
Glykolipidit ovat monimutkaisia lipidejä, jotka sisältävät hiilihydraattiryhmiä molekyylissä (yleensä D-galaktoosijäännös). Glykolipideillä on olennainen rooli biologisten kalvojen toiminnassa. Niitä löytyy pääasiassa aivokudoksesta, mutta niitä löytyy myös verisoluista ja muista kudoksista. Glykolipidejä on kolme pääryhmää: serebrosidit, sulfatidit ja gangliosidit.
Cerebrosidit eivät sisällä fosforihappoa tai koliinia. Ne sisältävät heksoosia (yleensä D-galaktoosia), joka on liitetty esterisidoksella aminoalkoholin sfingosiinin hydroksyyliryhmään. Lisäksi Cerebroside sisältää rasvahappoa. Näistä rasvahapoista yleisimpiä ovat lignoseriini-, nervoni- ja serebronihapot eli 24 hiiliatomia sisältävät rasvahapot. Serebrosidien rakenne voidaan esittää seuraavalla kaaviolla;
Serebrosidien tutkituimmat edustajat ovat nervonihappoa sisältävä nervon, aivohappoa sisältävä serebron ja glynosyyrihappoa sisältävä keratsiini. Serebrosidien pitoisuus on erityisen korkea hermosolujen kalvoissa (myeliinivaipassa).
Gangliosidit Kun gangliosideja hydrolysoituu, löytyy korkeampia rasvahappoja, alkoholisfingosiinia, D-glukoosia ja D-galaktoosia sekä aminosokerijohdannaisia: N-asetyyliglukosamiinia ja N-asetyylineuramiinihappoa. Jälkimmäinen syntetisoidaan kehossa glukosamiinista ja sillä on seuraava kaava:
Rakenteellisesti gangliosidit ovat suurelta osin samanlaisia kuin serebrosidit, ainoa ero on, että ne sisältävät yhden galaktoositähteen sijaan monimutkaisen oligosakkaridin. Yksi yksinkertaisimmista gangliosideista on punasolujen stromasta eristetty hematosidi:
Toisin kuin serebrosidit ja sulfatidit, gangliosideja löytyy pääasiassa aivojen harmaasta aineesta ja ne ovat keskittyneet hermo- ja gliasolujen plasmakalvoihin.
Kaikkia edellä käsiteltyjä lipidejä kutsutaan yleensä saippuoituneiksi, koska niiden hydrolyysi tuottaa saippuoita.
Sfingolipidien biosynteesi
Sfingolipidejä voidaan syntetisoida muista yhdisteistä. Niiden synteesi vaatii ennen kaikkea sfingosiinia, jota muodostuu palmitoyyli-CoA:sta ja seriinistä useiden peräkkäisten reaktioiden aikana; aktivoituja rasvahappoja tarvitaan asyyli-CoA-johdannaisten muodossa; tarvitaan myös
tai aktivoitu koliini CDP-koliinin muodossa sfingomyeliinien synteesiä varten tai hiilihydraattiluonteiset aktivoidut monomeerit niiden UDP-johdannaisten muodossa serebrosidien tai gangliosidien synteesiä varten.
Biologinen rooli
I. osallistuminen immuunijärjestelmään
a) Sfingolipidien aineenvaihdunta immuunijärjestelmän soluissa ja sekundaaristen lipidien välittäjien - keramidin, sfingosiinin, sfingosiini-1-fosfaatin ja keramidi-1-fosfaatin - muodostuminen ovat osa yhtä signaalijärjestelmää, joka ohjaa kypsymistä, erilaistumista lymfosyyttien aktivaatio ja proliferaatio vasteena antigeeneille ja mitogeeneille ja ohjelmoitu solukuolema efektoritoiminnan jälkeen.
b) Sfingomyeliinisyklin tuotteet sekä keramidisyntaasin estäjä - fumonisiini B1 - vaikuttavat T-lymfosyyttien pinta-antigeenien - CD3, CD4, CD8, CD25, CD45 - ilmentymiseen, muuttavat lymfosyyttien alapopulaatioiden välistä tasapainoa, estävät DNA-synteesiä kateenkorvan ja pernan normaaleissa soluissa ja proliferatiivisessa vasteessa mitogeeneille ja tukahduttaa immuunivasteen kehittyminen T-riippuvaisille antigeeneille in vivo.
Primaarisen immuunivasteen varhaisille vaiheille on tunnusomaista spesifisten prekursorien lisääntyminen lymfoidikudoksen spesifisessä mikroympäristössä, erilaistuminen efektorilymfosyyteiksi ja migraatio lymfoidisista elimistä vereen ja kudoksiin. Erityisesti T-lymfosyyttien migraatio riippuu antigeenin jakautumisesta ei-lymfoidisiin elimiin ja lymfosyyttien paikallisesta aktivaatiosta mononukleaaristen järjestelmien molekyylien toimesta.
c) Vaikuttaa adheesiomolekyylien ja MHC:n ilmentymiseen, samoin kuin solumigraatiotekijät säätelevät aktivoitujen lymfosyyttien suunnattua liikettä kudoksessa. Kaikentyyppisten efektorisolujen vuorovaikutus johtaa vieraan antigeenin poistoon kehosta. Sfingolipidien toiminta toteutuu TCR/CD3-kompleksin signalointireiteille ja sfingomyeliinisyklille yhteisten kohteiden tasolla. Sfingolipidit ovat immuunijärjestelmän tärkein ja korvaamaton osa, ja sen seurauksena tärkeä osa koko kehoa.
II- Osallistuminen solukalvojen rakenteeseen ja toimintaan.
Sfingolipidejä löytyy eläin- ja kasvisolujen kalvoista; ne ovat pulpaalisten hermojen ja aivojen lipidien myeliinivaipan pääkomponentti. Niitä ei juuri löydy rasvakerrostumasta.
Sovellus lääketieteessä
Sfingolipidejä käytetään syövän hoitoon. Monentyyppisiä kasvainsoluja ja kasvaimia voidaan tuhota hoidoilla, jotka lisäävät sfingolipidikeramidin pitoisuutta. On monia tapoja lisätä sfingolipidikeramidin määrää kasvaimessa, mutta niiden käyttöä vaikeuttaa se, että sfingolipidikeramidilla on keskeinen rooli solujen homeostaasissa: se metaboloituu helposti muiksi sfingolipideiksi, jotka edistävät kasvaimen kasvua, etäpesäkkeitä ja vastustaa potilaan immuunijärjestelmää. Tarve estää tällainen metabolinen konversio cfingolipidikeramidin synteesiin osallistuvien entsyymien samanaikaisen aktivoitumisen taustalla on kuvattu entsyymejä, jotka tulisi aktivoida tai estää, sekä lääkkeitä, metaboliitteja ja ravinnon komponentteja, jotka modifioivat kutakin entsyymiä. Allyylialkoholiryhmän merkitys keramidisfingolipidimolekyylissä ja useissa kasvainten vastaisissa aineissa korostetaan, ja osoitetaan, että hydroksyyliryhmä osallistuu fosfaatin siirtymiseen proteiinista proteiiniin muodostamalla fosfaattiesterin. Allyylihydroksyyliryhmä voi myös vähentää ketonien määrää mitokondrioiden ubikinoneissa tuottaakseen reaktiivisia happilajeja. Kfingolipidikeramidin tasoa kasvaimissa voidaan lisätä antamalla suoraan cfingolipidikeramidia tai sen analogeja; cfingolipidikeramidin muodostumisen stimulointi sen prekursoreista; sfingomyeliinin hydrolyysillä tai glykosfingolipidien hydrolyysillä; sfingosiinin asylointi. Lisäksi cfingolipidikeramidin suurempi pitoisuus voi johtua sen hitaamasta konversiosta sfingomyeliiniksi.
Keramidit – yksinkertaisin sfingolipidityyppi, joka koostuu sfingosiinista (tai joistakin sen johdannaisista) ja rasvahaposta (ne ovat tärkeä lipidikomponentti solukalvossa)
Sfingomyeliinin kaava:
Sfingomyeliini on eräänlainen sfingolipidi, jota löytyy eläinten solukalvosta. Hermosolujen aksonien myeliinivaippa on erityisen runsaasti tätä fosfolipidiä.
Sfingomyeliini on ainoa ihmisen fosfolipidi, jonka rungossa ei ole glyseroliosaa. Sfingomyeliini koostuu sfingosiinista, joka on liitetty esterisidoksella polaariseen ryhmään. Polaarinen ryhmä voi olla fosfokoliini tai fosfoetanoliamiini. Rasvahappo on kiinnittynyt sfingosiinin toiseen hiileen amidisidoksella.

2.Asetonin muodostumisreaktio.
Asetoni- orgaaninen aine, jonka kaava on CH3-C(O)-CH3, tyydyttyneiden ketonien yksinkertaisin edustaja.
Asetonia, joka muodostuu asetoasetaatin ei-entsymaattisessa dekarboksylaatiossa, ei käytetä kehossa. Se erittyy uloshengitysilmaan, hikirauhasten eritteisiin ja virtsaan. Normaalisti asetonin pitoisuus veressä on alhainen, eikä sitä määritetä tavallisilla reaktioilla.
Ketonikappaleet syntetisoituvat maksassa, kulkevat helposti mitokondrioiden ja solukalvojen läpi ja pääsevät vereen. Ne kulkeutuvat veren mukana kaikkiin muihin kudoksiin. Vain asetoasetaattia ja beeta-hydroksibutyraattia käytetään.


3. Esitä yleinen kuvaus asyylisyntetaasin rakenteesta ja sen aktiivisista keskuksista.
Rasvahapposynteesin reaktiot tämän entsyymin osallistuessa.
Tyydyttyneiden rasvahappojen biosynteesiin osallistuu kaksi entsyymikompleksia: asetyyli-CoA-karboksylaasi ja asyylisyntetaasi.
FA-syntetaasi sisältää 7 aktiivista keskusta.
Asyylisyntetaasi-multientsyymikompleksi sisältää asyylinsiirtoproteiini (ATP) eräänlaisena ytimenä, aktiivinen keskus on edustettuna fosfopantoteiini. Muita kompleksin entsyymejä ovat β-ketoasyylisyntetaasi (KS)– asyylisyntetaasin suurin domeeni (N-pää), jonka entsymaattinen aktiivisuus on koko prosessin ainoa palautumaton reaktio, asyylitransferaasi (AT)– siirtää happaman jäännöksen Acyl-CoA:sta ACP-alueen pantoteiiniosan HS-ryhmään, β-ketoasyylireduktaasi (KR), SISÄÄN- Hydroksiasyylidehydrataasi (HD), enoyylireduktaasi (ER) Ja asyylitransasetylaasi (AT).





Tämän jälkeen asyyli-ACP siirtyy uuteen synteesisykliin. Uusi malonyyli-CoA-molekyyli lisätään asyylinsiirtoproteiinin vapaaseen SH-ryhmään. Sitten asyylijäännös eliminoidaan ja se siirretään malonyylijäännökseen samanaikaisesti dekarboksyloimalla, ja reaktiosykli toistetaan.
Siten tulevan rasvahapon hiilivetyketju kasvaa vähitellen (jokaisessa syklissä - kahdella hiiliatomilla). Tämä tapahtuu, kunnes se pitenee 16 hiiliatomiin (palmitiinihapon synteesin tapauksessa) tai enemmän (muiden rasvahappojen synteesi). Tämän jälkeen tapahtuu tiolyysi ja muodostuu rasvahapon aktiivinen muoto, asyyli-CoA.